Fra DNA til protein - transkripsjon og translasjon
- Transkripsjon - fra DNA til budbringer RNA (mRNA)
- Fra mRNA til protein
- Regulering og uttrykk av gener hos eukaryoter
- Forskjellige typer RNA polymerase
- RNA prosessering
- Transkripsjonsfaktorer
- Genekspresjon hos prokaryoter
- Syntese av protein
DNA fungerer som templat for transkripsjon og inneholder oppskriften for oversettelse fra basesekvensen i DNA i kromosomer til aminosyrerekkefølgen i protein. Det er bare en av DNA-trådene som brukes som oppskrift i transkripsjonen, den kodende tråden (senstråden).
Proteinsyntesen skjer i to trinn. Først transkripsjon av DNA til mRNA og deretter translasjon av mRNA til protein. Transkripsjonen skjer i cellekjernen hvor det lages en en nøyaktig kopi av DNA i form av en enkelttrådet budbringer RNA (mRNA), men hvor thymin er erstattet av uracil. RNA skilles også fra DNA ved å ha ribose som sukker istedet for deoksyribose. RNA er mye mer ustabil enn DNA. RNase som bryter ned RNA er et allestedsnærværende enzym.
![]()
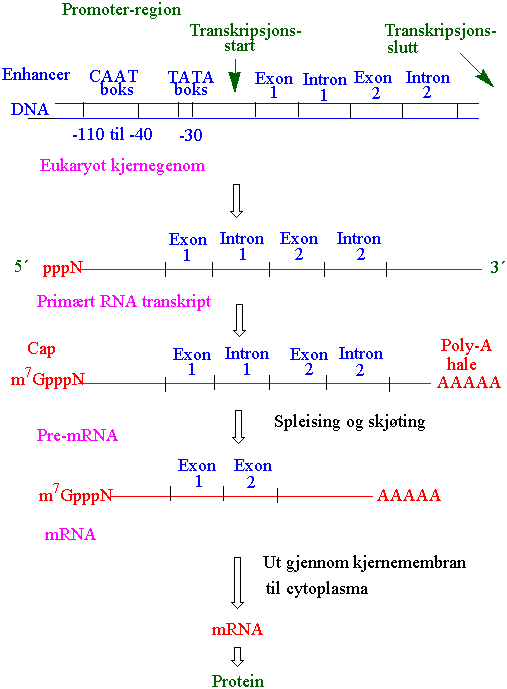
DNA sekvensen som koder for et protein blir transkriberet til et komplementært RNA transkript ved hjelp av enzymet RNA polymerase. Det primære RNA transkriptet må omdannes til en funksjonell mRNA som deretter fraktes ut av cellekjernen til cytoplasma hvor proteinsyntesen foregår. Den kodende sekvensen til et gen bestemmer rekkefølgen av aminosyrer i proteiner. Den kodende sekvensen hos de fleste eukaryoter avbrytes av ikke- kodende sekvenser kalt introner. Den kodende delen kalles eksoner. Ved transkripsjonen blir introns og exons transkribert slik at det lages et kontinuerlig RNA molekyl kalt primært transkript, som er mye større enn den endelige formen av mRNA. Grensen mellom introns og exons er konservert. Alle eukaryote introns begynner med GT og avslutter med AG (GT- AG regel). Hvert intron inneholder også en fast TACTAAC-boks. Exonene spleises og intronene fjernes i spleisosomer. Introns kuttes ut enzymatisk av pre-mRNA. Pseudogener er uvirksomme gener laget ved genduplisering eller mutasjon. Genduplisering kan gi genfamilier. Et vanlig gen inneholder sekvenser både oppstøms ved 5´-enden og nedstrøms ved 3´-enden. Disse regionene inneholder spesielle signaler som angir når og hvordan et gen skal transkriberes og translateres. Oppstrømsdelen kalles promoter.
Transkripsjon - fra DNA til budbringer RNA (mRNA)
Transkripsjonen begynner når RNA polymerase binder seg til promotersekvensen til genet. Bindingen av polymerase gir separering av de to trådene i DNA heliksen. En av trådene fungerer som templat og bestemmer rekkefølgen av basene i RNA komplementært til DNA. RNA polymerase kobler sammen RNA basene med en fosfodiesterbinding. Etterhvert som polymerasen beveger seg bortover DNA tråden åpner heliksen seg og RNA tråden forlenges. Transkripsjonen resulterer i en budbringerRNA (mRNA) som inneholder den proteinkodende sekvensen. Transkripsjonen skjer i 5´til 3´-retning. Ved 5´-enden av mRNA er det baser som er viktige for translasjonen, men som ikke koder for genproduktets aminosyresekvens. Dette er den 5´-ikketranslaterte delen. Et bestemt antall baser ved slutten av den kodende sekvensen utgjør 3´-ikketranslatert ende. Helt i starten på 5´-enden er det en cap ende som inneholder 7- metylguanosin bundet til den 5´-terminale nukleotidet ved en trifosfatgruppe. Cap-strukturen er nødvendig for at ribosomet skal kjenne igjen og binde seg til mRNA. Ribosomet bevege seg bortetter mRNA molekylet, men vil ikke starte translasjonen til protein før den treffer på et startkodon (AUG). Translasjonen vil deretter foregå inntil ribosomet treffer på et stoppkodon UAA, UAG eller UGA, hvor ribosomet frigis. Transkripsjon av gener hos eukaryoter katalyseres av RNA-polymerase II. Imidlertid deltar RNA polymerase I i syntesen av ribosomalt RNA (rRNA) og RNA polymerase III lager overføringsRNA (tRNA) (tranferRNA) og små nukleære og cellulære RNA molekyler. Bakterier bruker bare en type RNA polymerase i transkripsjonen.
Fra mRNA til protein
Basene i mRNA avleses på ribosomer i grupper på tre og tre kalt kodoner. Sammenhengen mellom kodoner og aminosyrer kalles genetisk kode. Det er 64 muligheter for kombinasjoner av tripletter av 4 baser (den genetiske koden), som i teorien kunne gi kode for 64 aminosyrer. Siden det bare er 20 aminosyrer som trenger kode har hver aminosyre flere mulige koder. I tillegg virker ett kodon som sluttsignal og ett kodon som startsignal for proteinsyntesen. Den genetiske koden uttrykkes vanligvis etter kodonene i mRNA. Seks kodoner koder for aminosyrene leucin, serin og arginin. Fire koder for glycin, valin, prolin og threonin. Methionin og tryptofan har ett kodon. AUG koder for methionin, men også for starten av leserammen for et protein. Translasjon av mRNA til protein starter alltid med AUG, altså methionin. Tre av kodonene er stoppkodoner: UAG (amberkodon), UGA (opalkodon) og UAA (ochrekodon). GUG og UUG er ofte startkodon hos bakterier.
På ribosomene (rRNA) skjer translasjonen ved hjelp av tRNA. Det er 61 forskjellige tRNA som har en gjenkjennelse sekvens komplementær til tripletten på mRNA. Protein består av en sekvens av aminosyrer. Det finnes varianter av samme protein og fenomenet kalles protein polymorfi. Et cistron koder for et polypeptid. Et protein kan bestå av fra 100 til 1000 aminosyrer og være sammensatt av subenheter. Ofte skjer det en modifisering av proteinet når det er ferdiglaget hvor andre typer molekyler hektes på.
Regulering og uttrykk av gener hos eukaryoter
Uttrykk av gener involverer interaksjon mellom DNA, mRNA, protein og RNA. Regulering av genekspresjon kan skje ved regulering av:
- 1) Transkripsjonshastighet.
- 2) Prosessering av pre-mRNA.
- 3) Transport av mRNA fra cellekjerne til cytoplasma.
- 4) Stabilitet og levetid for mRNA.
Gener som lager protein, tRNA og rRNA kalles strukturgener. Gener som kontrollerer når gener skal uttrykkes kalles regulatorgener. Noen kalles husholdsgener som uttrykkes i alle celler. Andre gener er vevsspesifikke og uttrykkes bare i en celle- eller vevstype. Kodende gener har flanker som koder for av og på, og hvor mye genprodukt som skal lages.
Reguleringen er kompleks og består av en interaksjon mellom ulike regioner av DNA, og binding av strukturproteiner og transkripsjonsfaktorer. Flankene til gener inneholder sekvenser som kan binde proteiner og finregulere transkripsjonen.
Hos bakterier (prokaryoter) kan flere gener være under kontroll en en promoter kalt operon. Operon finnes også hos arkebakteriene (Archaea), men ikke hos eukaryotene. Hvert eukaryot gen reguleres av sin egen promoter. DNA bindende proteinene er derfor sannsynligvis lite spesifikke slik at de kan aktivere eller undertrykke (repressere) en gruppe av relaterte gener. Regulering av genaktivitet er ofte et spørsmål om å regulere bindingen av RNA polymerase til promoteren, dvs. hvor tilgjengelig er kromatinstrukturen. Eukarotene har 3 forskjellige RNA polymeraser som transkriberer forskjellige klasser av gener. RNA polymerase binder seg ikke uten at en eller flere transkripsjonsfaktorer er tilstede. Hvis et repressor-protein binder seg til en spesiell del av DNA sekvensen i eller nær promotersekvensen vil bindingen av RNA polymerase blokkeres, og genet skrus av. I andre tilfeller vil forskjellige reguleringsproteiner binde seg til repressoren og løsne på affiniteten til promoteren, og genet skrus på. Positiv kontroll skjer ved at en transkripsjonsfaktor må binde seg først i eller nær promoteren før RNA polymerase bindes. Muligheten for at en transkripsjonsfaktor skal kunne binde seg til et kontrollelement og derved regulere genaktiviteten kan skje ved at mindre metabolitter som hormoner binder seg til transkripsjonsfaktoren, eller at transkripsjonsfaktoren modifiseres.
I promoterregionen (5´-oppstrømsregionen) er det for de fleste eukaryote gener flere sekevenser, kalt regulatorelementer, som er bindingssete for spesifikke DNA-bindende proteiner som er nødvendige for transkripsjonen. Promoterregionen står for regulering og start av transkripsjonen. Transkripsjonsfaktorer er proteiner som bindes til en spesifikk 5- 10 basepar DNA sekvens. Et gen kan inneholde flere sekvenser som vil modifisere ekspresjonen. Disse kalles "enhancere" hvis de stimulerer transkripsjonen eller "silencere" hvis de undertrykker transkripsjonen. Enhancere finnes ofte lenger oppstrøms enn promoterregionen. I de fleste tilfeller er det flere faktorer som virker sammen og påvirker transkripsjonen. Promoterregionen kan inneholde en TATA-boks og CAAT-boks. TATA- boksen er lokalisert ca. 10 basepar (bp) oppstrøms før transkripsjonsstart hos bakterier, og 15- 40 bp før start hos eukaryotene. TATA-boksen er bindingsstedet for transkripsjonskomplekset (RNA polymerase med transkripsjonsfaktorer). Eksempel på basesekvens i en TATA-boksen er TCACTATATATAG. Noen gener mangler TATA-boks, spesielt de som lager husholdsgener, f.eks. enzymer i Krebs syklus (konstitutive gener). Disse har istedet GC rike regioner (GC-boks) ca. 110 bp før transkripsjonsstart som regulerer transkripsjonen. Shine-Dalgarnoboks er et ribosombindende sete med konsensus AGGAGG ca. 7 bp før startkodon (ATG i bakterier). CAAT-boks ligger ca. 80 bp før transkripsjonsstart. Sekvensen AGGA kan erstatte CAAT. Både GC-boks og CAAT-boks er bindingsseterfor transkripssjonsfaktorer. DNA-sekvensen i disse boksene kalles cis-virkende sekvenser eller distale reguleringssekvenser. Transkripsjonsfaktorene som binder seg til de cis-virkende sekvenser kalles trans-virkende faktorer fordi genene som lager dem finnes andre steder i genomet. Transkripsjonsfaktorer som aktiverer transkripsjonen kalles aktivatorer og de som hemmer transkripsjonen kalles repressorer. cis-virkende sekvenser som påvirkes av hormoner eller andre signalstoffer kalles responselementer.
Hos noen bakterier og virus kan den kodende sekvensen starte fra første andre eller tredje base i kodonet og derved bruke forskjellige leserammer og derved lage flere proteiner.
Forskjellige typer RNA polymerase
Det er 3 forskjellige typer av enzymet RNA polymerase hos eukaryoter:
1) RNA polymerase I lokalisert i nukleolus transkriberer gener som koder for store ribosomale RNA. En plantecelle kan inneholde ca. 10 millioner kjernekodete ribosomer (80 S). Gener som koder for ribosomalt RNA (rRNA) finnes gjentatt i tandem spredt rundt i genomet med fra 1000-10.000 kopier per haploid genom. Sammenkoblingen av ribosomalt RNA (rRNA) og ribosomale proteiner skjer i nukleolus i kjernen. Transkripsjonen av rDNA gir pre rRNA som inneholder 3 av de 4 rRNA som inngår i ribosomene, nemlig den lille subenheten 18S, 5.8S og den store subenheten 25S rRNA. Ved avkutting i 5´-enden av den lille subenheten og internt transkriberte spacere (ITS) lages ferdig rRNA. I tillegg skjer det modifikasjoner som 2´-O-ribose metylering og pseudouridylering. Det er en rekke små nukleolus RNA (snoRNA).
2) RNA polymerase II lager pre-mRNA i kjernen, UsnRNA i spleisosom og koder for cytoplasmatiske proteiner.
3) RNA polymerase III lager små cytoplasma RNA (scRNA) som 4.5 S og 5S ribosomale RNA (5S rRNA), U6sn RNA, og tRNA.
RNA polymerase hos eukaryoter er store molekyler med molekylvekt opptil 500 000kDa og består av 7-14 polypeptider. Hver av disse polymerasene kjenner igjen en spesiell DNA basesekvens eller gruppe av sekvenser, men RNA polymerase trenger hjelp av flere protein transkripsjonsfaktorer (TF). For de 3 polymerasene får disse navn TFI, TFII, TFIII etterfulgt av en bokstav siden det er flere av dem, f.eks. TFIIIH. De 3 RNA polymerasene har også egne promotersekvenser
RNA prosessering
RNA prosessering skjer i flere trinn. Siden det primære transkriptet (pre-mRNA) inneholder både exons og introns må introns fjernes. Før skjøting og fjerning av introns må pre-mRNA modifiseres på to måter:
1) Polyadenylering er påhenging av en lang rekke adenosin nukleotider i 3´-enden av transkriptet, et merke (tag) kalt poly-A-hale. Det er vanligvis 100-200 adenin som hektes på hos plantegener katalysert av poly-A-polymerase. Poly-A påsettingssignalet er sekvensen AATAAA som finnes i 3´-transkribert uoversatt region, 10-30 baser fra poly-A halen.
2) Påsetting av en hette (cap) bestående av 7-metylguanylat i 5´-enden av transkriptet. Poly- A-halen og cap i den andre (5´-cap) er nødvendig for at mRNA skal kunne passere kjerneporene. Poly-A-halen og 5´-cap beskytter mRNA mot nedbrytning av RNase og øker derved stabiliteten og levetiden til mRNA. Nylaget pre-mRNA pakkes i en proteinpartikken kalt heteronukleær ribonukleoproteinpartikkel (hnRNP partikkel). En slik partikkel kan inneholde små nukleære ribonukleoproteiner (snRNP) som inngår i et spleisosom kompleks. Til slutt skjer skjøtingen hvor intronene fjernes og exonene skjøtes sammen og gir den endelige funksjonelle mRNA. mRNA transporten ut av kjernen er regulert, og mRNA som har kommet ut i cytoplasma dirigeres mot endoplasmatisk retikulum. mRNA i cytoplasma kan nedbrytes etter bruk med større eller mindre hastighet, avhengig av hvilket protein som skal lages. I hvilken grad mRNA translateres til protein kan bestemmes av stabiliteten til mRNA, folding og sekundærstruktur av mRNA og i hvilken grad frekvensen av uvanlige kodoner er brukt.
Transkripsjonsfaktorer
Transkripsjonsfaktorene har generelt 3 strukturelle egenskaper. De har:
- 1) Et domene som binder spesifikt til et stykke av DNA.
- 2) Et transkripsjonsaktiverende domene.
- 3) Et ligandbindende domene.
Det finnes 5 forskjellige hovedtyper av transkripsjonsfaktorer basert på forskjellige DNA- bindende domener:
1) Leucin zippere har områder i proteinet med 4-7 stykker av aminosyren leucin som forekommer i regulære intervall i en a-heliks på 35 aminosyrer slik at de kommer på linje på samme side i heliksen. Dette gir en hydrofob stripe langs heliksen som vil reagere med en lignende struktur på et annet protein, slik at proteiner med leucinzippere danner lett en dimer. Transkripsjonsfaktorene AP-1 fra menneske og GCN4 fra gjær er eksempler på leucinzippere. Disse inneholder lik region av basiske aminosyrer i deres karboksyl-terminale region, som er det DNA-bindende domene TGACTCA. Fos og Jun er andre eksempler.
2) Basiske zippere er transkripsjonsfaktorer hvor andre hydrofobe aminosyrer kan erstatte leucin, f.eks. Opaque 2 som regulerer transkripsjonen av noen frølagringsproteiner i mais. Et annet eksempel er transkripsjonsfaktorer som binder abscisinsyre responselementer.
3) Heliks-løkke-heliks dimer ("helix-loop-helix"). Disse er lik leucin zippere unntatt at hver monomer inneholder 2 a-heliks domener, atskilt ved en region hvor proteinet kan lage en løkke utover. F.eks. GTP bindende protein i fytokromregulerte gener.
4) Helix-vending-helix ("helix-turn-helix")) To a-helixer atskilt av en vending. Heliks- vending-heliks transkripsjonsfaktorer er kjent for å regulere biosyntesen av anthocyaniner.
5) Zinkfinger. Steroidhormon receptorer er potensielle DNA bindende proteiner. De har en region som er rik på basiske aminosyrer og en region som er rik på aminosyren cystein. Den cysteinrike regionen kan reagere med zink og danne en zinkfinger. Eksempel er COP1 i Arabidopsis.
Små RNA
Betydningen av små RNA , små interfererende RNA (siRNA), mikro-RNA (miRNA) ble oppdaget i studiet av transgene petunia (Petunia hybrida) hvor man ønsket å se virkningen på blomsterfargen ved å sette inn både sens- og antisens-transkript for chalkon syntase, et sentralt enzym i biosyntesen av anthocyaniner. Sens- og antisenstranskriptene var likt med mRNA transkriptene som ble laget endogent i planten. Man tenkte at antisenstranskriptet skulle binde seg til endogent mRNA via baseparring og derved hindre translasjon. Men det ble også dannet mindre pigment fra sens-transkriptet. Det skjedde en kosuppresjon, det innsatte eksogene genet undertrykte det endogene, via en posttranskripsjonsregulering via RNA-styrt DNA-metylering, men også transkripsjonsslukking via DNA-metylering.
Mkro-RNA (miRNA)blir laget fra MIR-gener, en type ikke proteinkodende gener som enzymet RNA polymerase II transkriberer til enkelttrådet miRNA, men primær mikro-RNA har en hårnålsløkke som folder miRNA omkring seg selv slik at det blir dannet dobbelttrådet RNA (dsRNA). Det finnes fler hundre forskjellige MIR-gener. Mikro-RNA kan binde til til messenger-RNA (mRNA, budbringerRNA) som har komplementær sekvenslikhet, og påvirker derved stabiliteten til mRNA via posttranslasjonsgenslukking. Dobbelttrådet RNA blir delt i lengder på 21-24 basepar via en endoribonuklease RNase kalt DICER og DCL (DICER-lignende). Blir O-metylert i 3-terminal ende for å hindre uridylering og nedbrytning. Planter kan lage ferdig miRNA via DICER, mens dyr trenger to enzymer kalt Drosha og Pasha i kjernen, og DICER i cytoplasma. Drosha er en klasse 2 ribonuklease III, og Pasha er et RNA-bindende protein.
Små interfererende RNA (siRNA) blir laget av enzymet RNAavhengig RNA polymerase som lager dobbelttrådet RNA (dsRNA) eller enkelttrådet hårnols RNA. Slike små interferernde RNA deltar i forsvar mot virus via transkripsjonsgenslukking, og gir resitens ved å ha små RNA med komplementær basesekvens som virus-RNA.
Det er også blitt funnet fasete små interfererende RNA (phasiRNA) som deltar i genregulering, og kodet av en type gener hos planter kalt phasi-RNA produserende gener, PHAS-gener. En type av disse er transvirkende små interfererende RNA (tasiRNA).
Små RNA binder seg til ARGONAUT-protein (AGO) som en del av det RNA-induserte slukningskomplekset RISC, som kan binde og modifisere DNA eller RNA som gir slukking av avskruing av gener. AGO har fått navn ett blekkspruten Argonauta. Det finnes mange forskjellige AGO som deltar i forsvar mot RNA-virus og gir RNA-styrt DNA-metylering. Transposoner holdes i sjakk via DNA-metylering. Små interferernde RNA (siRNA) kan fraktes til andre celler, for eksempel siRNA fra den vegetative cellen i pollenkorn fraktes til den generative cellen hvor den gir slukning av transposoner. Dette gir epigenetisk styrking av transposoner, ofte GC-metylerte, og repeterte sekvenser.
Mikro-RNA (miRNA) deltar i bestemmelsen av hva som skal bli overside og underside på et blad, påvirker bladranden på blad, og deltar i dannelse av siderøtter og adventivrøtter.
Lysregulering av gener
Lys styrer en rekke utviklingstrekk hos plantene kalt fotomorfogenese. Den lille subenheten til rubisko (rbcS) og klorofyll a/b bindende protein (cab) blir transkribert på lavt nivå i etiolerte planter og transkripsjonen øker 20-50 ganger i lys. Disse genene har et oppstrøms lysresponderende element.
Multigenfamilier
Hos plantene er det gener som finnes i så stort antall at de utgjør multigenfamilier. Genantallet kan forandres i en art ved genduplisering hvor det lages kopier av gener. Dette kan også skape nye gener med nye funksjoner. Genduplisering kan skje på forskjellige måter. En måte er ulik overkrysning. Det kan også skje ved polyploidi. Genduplisering kan også skje ved transposisjon hvor et stykke av en DNA sekvens på et sted kopieres og settes inn i en ny posisjon på et annet sted. Dette gir økt toleranse for mutasjoner. Eksempler på genfamilier er gener som koder for lagringsproteiner i frø. Den lille subenheten til det CO2-fikserende enzymet i fotosyntesen, leghemoglobin og nodulin som innngår i nitrogenfikseringen, samt enzymet chalcon syntase i syntesen av flavonoider er andre eksempler.
Betydningen av genduplisering med Hox-familien er godt illustrert ved utvikling av kroppsplanet hos dyr. Andre multigenfamilier er repetert side ved side slik at mange gener med samme funksjon ligger ved siden av hverandre i tandem. Dette skjer hvis det er stort behov for genproduktene f.eks. syntese av rRNA.
Genekspresjon hos prokaryoter
Prokaryoter inneholder mindre DNA enn eukaryotene og DNA er tynt dekket med protein som deltar i folding av DNA. Noen av proteinene trengs til reparasjon, rekombinasjon og transkripsjon. Genreguleringen er mindre kompleks hos prokaryoter enn eukaryoter. Hos prokaryotene er genene samlet i operoner. Et operon er en samling strukturgener og reguleringssekvenser. Det meste kjente eksemplet er det induserbare laktoseoperonet hos tarmbakterien Escherichia coli, studert av Jacob og Monod på 1960-tallet. lac-operonet lager enzymene som er nødvendig for å omsette disakkaridet laktose. Det er 3 strukturgener i operonet: 1) s (sigma)-faktorerb-galaktosidase som spalter laktose til glukose og galaktose. 2) permease som er et bærerprotein som frakter laktose inn i cellen og 3) en transacetylase med ukjent frunksjon. Et oppstrøms regulatorgen lager et repressorprotein som kjenner igjen og binder seg til en nukleotidsekvens kalt operator. Mellom regulatorsekvensen og operator ligger en promoter som binder RNA polymerase og hvor transkripsjonen starter. Starten skjer på 5´-enden av genet, men RNA polymerase transkriberer fra 3´-enden til 5´-enden på den motsatte tråd. Dette er en konvensjon slik at sekvensen til mRNA skal matche DNA sekvensen til genet. RNA polymerase hos prokaryoter inneholder polypeptider kalt som er nødvendig for transkripsjonen. Hvis ikke laktose er tilstede så danner repressor et kompleks med operator og hindrer transkripsjon. Hvis laktose er tilstede endres konformasjon av repressor og transkripsjonen starter. Lac-operonet kan også reguleres av glukose slik at hvis cellene får glukose sammen med laktose undertrykkes uttrykket av lac-operonet. Glukose gir effekt ved å senke konsentrasjon av syklisk AMP (cAMP). cAMP binder seg til et aktivatorprotein (katabolitt aktivator protein).
Genregulering hos gjær
De fleste genene hos gjær kodes av oppstrømsaktiverende sekvenser (OAS). GAL4 koder for en transkripsjonsfaktor som kan binde seg til OAS ca. 200 basepar oppstrøms for 6 forskjellige gener som deltar i metabolismen av galaktose og aktivere alle disse samtidig. Når et GAL80 protein bindes til GAL4 protein stoppes transkripsjonen
Syntese av protein
Protein er en sekvens av aminosyrer bundet sammen i peptidbinding, etter oppskrift fra den genetiske koden i basesekvensen i mRNA ("messenger-RNA", budbringer-RNA). -karboksylsyrekarbon fra en aminosyre henger sammen med -nitrogen i den neste aminosyren i et polypeptid. Den lineære polypeptidkjeden kalles primærsekvens. Polypetidkjeden danner mer komplekse sekundærstrukturer kalt -heliks og -plateark, som foldes til en mer stabil tertiærstruktur. To eller flere polypeptidkjeder kan settes sammen og danne kvartærstrukturer. Mesteparten av proteinet lages i cytoplasma, og noe i plastider og mitokondrier. Plastider og mitokondrier er rester etter endosymbionter og har proteinsyntese som ligner på det man finner i bakterier. For hvert kodon i translasjonen fra mRNA finnes det et tilsvarende antikodon på tRNA (transfer-RNA). Det finnes 31 forskjellige tRNA, hver bundet til en aminosyrer i 3´-enden. tRNA, med opptil 90 baser, er et molekyl med 3 løkker: D-, T- og antikodon-løkken. Løkkene kan inneholde uvanlige ribonukleotider. Siden den genetiske koden er slik at en aminosyrer kan ha flere kodoner, trengs det færre tRNA enn anntall kodoner. Aminoacetyl-tRNA-syntetase er enzymer som fester aminosyrene til tRNA i en ATP-avhengig totrinnsprosess. Først lager enzymet et aminoacyladenylat og deretter festes den aktiverte aminosyren til hydroksylgruppen i 3´-enden på tRNA. Ribosomene, 70S i mitokondrier og plastider og 80S i cytosol, er satt sammen av to subenheter. (S er Svedberg- enheter som et mål på sedimentering i ultrasentrifuge i sukrose-tetthetsgradienter). Ribosomene, som inneholder RNA og opptil 80 forskjellige proteiner, styrer sammenkoblingen av aminosyrer. mRNA og ribosomene beveger seg i forhold til hverandre ettersom proteinsyntesen pågår. Ribosomene kan være frie eller bundet til membraner. Proteinsyntesen er delt i 3 faser: initsiering, forlengelse (elongering) og terminering. Startkodonet AUG, som koder for en spesiell initsiering-tRNA festet til methionin, bestemmer den videre leserammen av nukleotidsekvensen i mRNA, avlest i 5´- til 3´- retning. Derfor starter alle proteinene med methionin, men startmethionin kan eventuelt fjernes etterpå av en aminopeptidase som blottlegger den neste aminosyren i sekvensen. Flere ribosomer er festet til mRNA samtidig, kalt polyribosomer eller polysomer. Initsieringen av proteinsyntesen hos eukaryoter er kompleks. Den trenger energi i form av GTP, og flere initsieringssfaktorer (elF1 til eIF6) før met-tRNA bindes til 5´-cap på mRNA og den lille 40S subenheten i 80S ribosomet. Til slutt bindes 60S subenheten i ribosomet og syntesen av en polypeptidkjede starter. I motsetning til prokaryoter er mRNA hos eukaryoter vanligvis monocistronisk og koder for bare en type protein. Den sekvensielle forlengelsen av proteinkjeden ved påkobling av nye aminosyrer skjer på 3 seter på 80S ribosomet: peptidyl- tRNA bindingssete (P-sete), Aminoacyl-tRNA bindingssete (A-sete) hvor neste kodon blir avlest og Exit-sete (E-sete) hvor tRNA etter å ha avgitt sin aminosyre bindes et kort øyeblikk før den fjernes. Forlengelsen av polypeptidkjeden fortsetter inntil ribosomet treffer et stoppkodon (UAA, UAG eller UGA). Den ferdige polypeptidkjeden blir frigitt fra ribosomet ved hjelp av flere frigivelsesfaktorer (eRF, "releasing factor") og GTP. Polysomer som er festet til endoplasmatisk retikulum (ER) styrer transporten av protein inn i lumen i ER. Proteiner som skal inn i plastider eller mitokondrier har med seg en ekstra aminosyresekvens, signalsekvens, som virker som merkelapp og leder proteinet fram til riktig sted i cellen. aaab
mRNA hos bakterier og plastider mangler cap, er polycistronisk og inneholder flere leserammer. 16S rRNA på den lille subenheten 30S i 70S ribosomet finner startkodonet AUG ved hjelp av en Shine-Dalgarno basesekvens oppstrøms for startkodonet. Polypeptider fra prokaryoter, plastider og mitokondrier starter med formylmethionin, men denne fjernes vanligvis etterpå av en deformylase. Polysomer som er bundet til thylakoidmembraner kan sette proteinet rett inn i membranen etterhvert som det lages.
Regulering av proteinsyntesen og etterbehandling
Syntesen av protein er strengt regulert av behovene i plantecellene styrt av indre forhold og ytre omgivelsesfaktorer. Syntesen av protein kan styres av tilgangen på initsieringsfaktorer. Initsieringsfaktor eIF2 binder GTP og blir etter start av proteinsyntesen frigitt som IF2-GDP. GDP må fosforyleres på nytt før den kan delta i proteinsyntesen og dette er eksempel på et trinn som kan styres. Etter at proteiner har nådd fram til bestemmelsesstedene i cellen blir de modifisert og etterbehandlet. Signalsekvenser fjernes proteolyttisk og proteinene blir foldet i en stabil 3-dimensjonal struktur. Hydrofile og hydrofobe sidekjeder på aminosyrene, ladninger og disulfidbindinger bestemmer konformasjonen.
Chaperoner er en gruppe proteiner som stabiliserer og deltar i folding av proteiner. Proteiner kan bli nedbrutt proteolyttisk av proteaser og nysyntetiseret kontinuerlig. Vakuolene inneholder mange proteaser. Levetiden til proteiner bestemmes av signalsekvenser i proteinet bl.a. i N-terminal ende. I cytoplasma og kjernen blir proteiner nedbrutt av proteasomer. Proteiner som skal nedbrytes av proteasomer blir merket med ubiquitin.