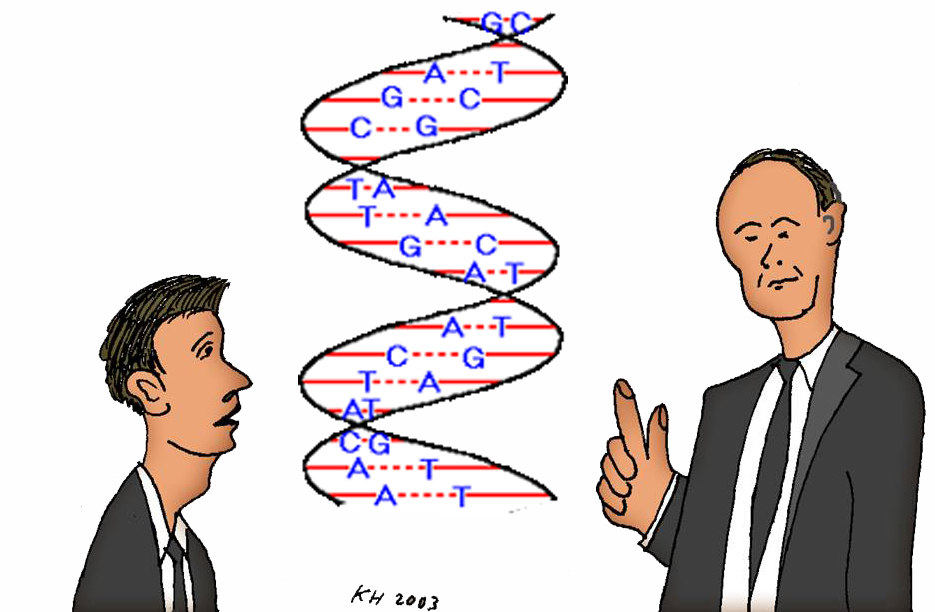
Nukleinsyrene ble oppdaget i 1869 av den sveitsiske kjemikeren Friedrich Miesner. Da de først ble påvist i cellekjernene, fikk de navnet etter nucleus som betyr kjerne. Tyskeren Albrecht Kossel spaltet i 1880-åra nukleinsyrene i byggesteinene fosforsyre og pentoser (en type sukker med fem karbonatomer), samt fem organiske baser. To av disse basene tilhører stoffgruppa puriner og ble kalt adenin og guanin. De tre andre er pyrimidiner og ble kalt cytosin, thymin og uracil. Den russisk-amerikanske kjemikeren Phoebus Aaron Theodor Levene fant at i nukleinsyremolekylet danner ett fosforsyremolekyl, ett pentosemolekyl og én av basene en tredelt enhet som han kalte et nukleotid. Selve nukleinsyremolekylet er dannet ved at fosforsyra i ett nukleotid er koplet til pentosegruppa i neste nukleotid. Slik blir det bygget opp en pentose-fosfat-ryggrad. Levene oppdaget dessuten at det fins to typer nukleinsyrer, avhengig om sukkeret er ribose eller deoksiribose. Den første ble kalt ribosenukleinsyre RNA og inneholder basene adenin A, guanin G, cytosin C og uracil U; den andre deoksiribosenukleinsyre DNA med basene adenin A, guanin G, cytosin C og thymin T. – Merk at uracil i RNA er byttet ut med thymin i DNA.
Etter Den 2. verdenskrig fant den østerrikske biokjemikeren Erwin Chargaff en merkelig egenskap hos DNA-molekylet. Etter møysommelige analyser konstaterte han at det var like mange adenin- og thyminmolekyler og like mange guanin- og cytosinmolekyler per DNA-molekyl. Og ikke nok med det: DNA fra visse organismer hadde et overskudd av adenin og thymin, mens andre arter hadde et overskudd av guanin og cytosin.
Men til å begynne med var det ingen som koplet nukleinsyrene til genene. Man trodde fullt og fast på at arvestoffet besto av proteiner, de mest kompliserte organiske forbindelsene man kjente til. Nukleinsyrene ble antatt å være en slags støttesubstans i cellekjernen. De var altfor enkelt oppbygd til å fungere som noe arvestoff. I høyden spilte de bare en birolle. Det er bare fire baser i nukleinsyrene, mens 21 aminosyrer bygger opp proteinene. Vi skulle derfor tro at proteinene inneholder mest informasjon.
Nådestøtet mot proteinhypotesen kom i 1952 (riktignok hadde tidligere forsøk med bakterier vist at nukleinsyrene nok var viktigst). Dette året utførte amerikanerne Alfred D. Hershey og Martha Chase et eksperiment hvor de lot virus som angriper bakterier, bakteriofager, ta opp radioaktivt merket svovel og fosfor. Svovelet bygges inn i proteinene som danner kappen til bakteriofagen, mens fosforet bygges inn i nukleinsyra DNA som fins inni bakteriofagen. Disse merkete bakteriofagene fikk infisere umerkete bakterier. Da viste det seg at kun det radioaktive fosforet trengte inn i bakteriene! Det radioaktive svovelet ble sittende igjen utapå. Av den radioaktive nukleinsyra ble det nye bakteriofager med nye proteinkapper. Disse hadde nukleinsyre med radioaktivt fosfor, men proteinkappe uten radioaktivt svovel. Konklusjon: Genet består utelukkende av nukleinsyre! – Men hvordan var strukturen til denne viktige forbindelsen ut over det man til da visste om pentose-fosfat-ryggraden og de organiske basene?
Så kom Watson og Crick på banen!
Amerikaneren James Dewey Watson karakteriserer seg sjøl som en person som levde i håpet om at genenes natur skulle bli avslørt uten at han behøvde å lære noe om kjemi. Da han studerte ved universitetet i Chicago, var han mest interessert i fugler og unngikk et hvert kurs i kjemi eller fysikk som forekom ham over middels vanskelighetsgrad. Etter utdannelsen i USA fikk han et forskningsstipend til København for å arbeide med bakteriofager og deres DNA. Oppholdet i København var ikke spesielt vellykket, og etter mye fram og tilbake kom han til Cambridge-universitetet i England hvor han i 1951 fikk kontakt med Francis Crick ved Cavendish-laboratoriet.
Om engelskmannen Francis Crick sa visse av kollegaene at han hadde snakket uavbrutt i 35 år uten at dette hadde resultert i noe som kunne sies å være av fundamental verdi. Crick hadde begynt på hovedfag da Den 2. verdenskrig brøt ut i 1939. Som de fleste britiske forskere stilte han seg til rådighet for forsvaret og arbeidet ved Admiralitetets forskningsapparat, hovedsakelig med å framstille magnetiske og akustiske miner. Han forlot Admiralitetet i 1947 og hadde samtidig mistet lysten på å studere fysikk. Han begynte istedenfor å studere biologi ved Cambridge-universitetet, først Strangeways-laboratoriet, siden, i 1949, flyttet han over til Cavendish-laboratoriet, med tanke på at han endelig ville prøve å ta doktorgrad. Han sluttet seg blant annet til biofysikeren Max Perutz som arbeidet med røntgenstrålenes diffraksjon i hemoglobinkrystaller, blodets røde fargestoff. Leder av Cavendish-laboratoriet var Sir Lawrence Bragg som hadde fått Nobelprisen for sin innsats for krystallografien. I nesten 40 år hadde han sett hvordan røntgendiffraksjonsmetodene hadde hjulpet med å klarlegge sjøl de mest innviklete strukturer. Crick ble i 1950 innrullert ved Caius College, hvor han skulle jobbe under ledelse av Max Perutz. I 1954 fikk han sin doktorgrad om røntgendiffraksjon på proteiner. Det er verd å nevne at før 1947 visste Crick ingen ting verken om biologi, organisk kjemi eller krystallografi.
Det var slett ikke meningen at Watson og Crick skulle jobbe med strukturen til DNA-molekylet. For det første skulle Watson egentlig fullføre sitt stipendium og studere bakteriofager i København. Noe sidesprang til Cambridge ville ikke stipendiatrådet vite noe av, ei heller anbefale. For det andre burde Crick bli ferdig med den planlagte doktorgraden. For det tredje gikk ryktene om at den berømte amerikanske kjemikeren ved Cal Tech, Linus Pauling, arbeidet med DNA-strukturen. Han hadde jo tidligere høstet ære ved avsløringen av proteinmolekylenes alfaheliks og burde være mer enn kompetent til å jobbe med andre kompliserte, organiske kjedemolekyler. For det fjerde – og det var viktigst – eksisterte det ei forskergruppe i London som allerede var godt i gang med den molekylære utforskningen av DNA, Maurice Wilkins og Rosalind Franklin ved King’s College. Den engelske sansen for fair play gjorde det utillatelig for andre å blande seg inn i deres problem. Særlig fordi Wilkins og Crick var jevngamle og kjente hverandre personlig.
Maurice Wilkins og Rosalind Franklin var begge ansatt som forskningsassistenter under James Randall ved King’s College. Wilkins – som opprinnelig var fra New Zealand – arbeidet med fysikk og brukte først og fremst røntgendiffraksjonsmetoder i forskningen sin. Han hadde jobbet med krystallinsk DNA i flere år. Rosalind Franklin, utdannet i kjemi ved Cambridge-universitetet i 1951, var ekspert på røntgenkrystallografi. Hun hadde allerede gjort fortjenestefulle studier av strukturen til grafitt og andre karbonforbindelser før hun ankom King’s College. Det var hun som laget de fleste røntgendiffraksjonsbildene av DNA. Det var hun som antok at pentose-fosfat-ryggraden ligger utapå DNA-molekylet, mens både Watson og Crick, og Pauling fremdeles trodde ryggraden var i sentrum. I 1952 tok hun også det avgjørende bildet av DNA i B-formen som viste at DNA-molekylet danner en spiral.
Trass de nevnte hindringene, fortsatte Watson og Crick å tenke på hvordan DNA-mysteriet kunne løses. Og mot Cavendish-laboratoriets ønsker begynte de å legge slagplaner hvordan de skulle komme i mål før kollegaene ved King’s College og Linus Pauling. Personkjemi er en viktig faktor her: Watson var full av ideer og pågangsmot overfor de ulike løsningene av problemet. Hans egen teoretiske bakgrunn var kanskje ikke på topp, men han hadde en fabelaktig evne til å suge til seg de siste forskningsresultatene, for eksempel de til Chargaff og Hershey og Chase, og bygge videre på dem. I tillegg hadde han en god porsjon intuisjon – for eksempel da han besluttet å bygge DNA-modellen med to istedenfor tre kjeder fordi biologiske objekter alltid opptrer parvis. Crick var utadvendt og så entusiastisk overfor det han i et hvert øyeblikk jobbet med at han drev sine kollegaer til vanvidd med høyrøstet prat. Dessuten var han atskillig bedre skolert i teoretisk fysikk enn Watson og kunne tolke og forstå sammenhengene som framkommer ved røntgendiffraksjonskrystallografi. Problemet til Watson og Crick var imidlertid at de ikke hadde tilgang til det empiriske DNA-materialet. Dette satt kollegaene ved King’s College på, i form av praktfulle røntgendiffraksjonsfotografier. Og samarbeidsviljen derfra var så som så. Særlig Rosalind Franklin var skeptisk. Hun var også en helt motsatt personlighetstype og mislikte sterkt de lettvinte løsningene og generaliseringene uten bakgrunn i harde fakta som Watson og Crick hele tida gjorde seg til talsmenn for. Hun betvilte for eksempel DNA-molekylets spiralstruktur helt til hun endelig hadde de ugjendrivelige røntgendiffraksjonsbevisene.
I februar 1953 kom sjokkmeldingen at Linus Pauling hadde funnet fram til løsningen på DNA-molekylet. Modellen var en spiral med tre kjeder og med pentose-fosfat-ryggraden i sentrum. Umiddelbart ville ingen bestride modellen til Pauling. Men Watson fant snart ut at noe ikke stemte. Fosfatgruppene i Paulings struktur var ikke ionisert, hver av dem inneholdt et bundet hydrogenatom og hadde dermed ingen nettoladning. Paulings nukleinsyre var ingen syre! – Og nukleinsyrene har klart sure egenskaper! – Enda verre var at de fatale hydrogenatomene inngikk i de hydrogenbindingene som holdt sammen de tre kjedene. Uten disse bindingene ville Paulings modell falle fra hverandre som et korthus. En kjempe som Linus Pauling hadde altså glemt sin elementære skolekjemi! – Historien forteller at Watson og Crick feiret Paulings nederlag med whisky på deres stampub, Eagle.

I 1952 tok Rosalind Franklin et av biologiens mest berømte bilder. Dette er det før omtalte røntgendiffraksjonsbildet som viser DNA i B-formen, dvs. den tredimensjonale strukturen molekylet har når det er omgitt av en stor vannmengde. Franklin viste dette bildet til Watson, som umiddelbart skjønte at DNA-molekylet danner en spiral. Både Wilkins og Franklin mente at basene måtte være i midten og ryggraden på yttersida. Etter å ha sett dette skjellsettende bildet begynte Watson å sysle med modeller med to kjeder med ryggraden ytterst og basene i sentrum. Problemet var basene. De opptrer jo i fire ulike typer og med to fundamentalt forskjellige størrelser: puriner og pyrimidiner. Så lenge de kunne plasseres på yttersida av spiralen, behøvde ingen å bekymre seg med dem. Hvis de derimot ble anbrakt i midten, så forelå nesten uoverstigelige problemer å få pakket sammen to eller flere kjeder med uregelmessige basesekvenser. Dessuten manglet Watson og Crick ei rekke viktige eksperimentelle data fra King’s College. Disse tilegnet de seg på en heller ufin måte! Det forholdt seg nemlig slik at Max Perutz var medlem av en komité oppnevnt av Det medisinske forskningsråd for å inspisere aktivitetene ved James Randalls laboratorium. Randall hadde derfor bedt sine underordnete, blant annet Wilkins og Franklin, om å lage omfattende oversikter over hva de hadde prestert. Perutz, som medlem av komiteen, hadde fått kopi av oversiktene. Så snart han fant avsnittene skrevet av Wilkins og Franklin, viste han dem til konkurrentene Watson og Crick. Det var imidlertid ikke meningen at de skulle anvende dem uten videre til sin egen forskning!
Ved hjelp av Wilkins og Franklins empiriske data kunne Watson og Crick starte på ny frisk igjen. Nå hadde de alle målingene som skulle til for å kunne justere den endelige DNA-modellen. Først forsøkte Watson en tokjedemodell hvor baseparene var koblet sammen etter likt med likt-prinsippet, dvs. adenin mot adenin, thymin mot thymin osv. Men dessverre, da den amerikanske krystallografen Jerry Donohue dagen etter konfronterte Watson med at han hadde anvendt de gale tautomere formene av basene – enol- istendefor keto-formen – måtte han innse at likt med likt-modellen falt like fullstendig i grus som Paulings trekjedemodell.
Etter likt med likt-nederlaget, startet Watson neste dag med nytt mot. Plutselig ble han oppmerksom på at et adenin/thymin-par som ble holdt sammen av to hydrogenbindinger, hadde samme form som et guanin/cytosin-par med minst to hydrogenbindinger mellom basene. Alle hydrogenbindingene så ut til å danne seg naturlig, det var ikke nødvendig med noen fiksfakserier for å gjøre de to typene basepar identiske i form! Han ante at de nå hadde svaret på gåten hvorfor antallet purinkomponenter alltid var nøyaktig likt antallet pyrimidinkomponenter. Dermed framsto Chargaffs observasjon som en konsekvens av at DNA har natur av en dobbeltspiral! Da Crick ble konfrontert med dette resultatet, varte det ikke lenger enn til lunsj før han flakset inn på puben Eagle og ropte at nå hadde de funnet livets hemmelighet!
Kollegaene ved King’s College aksepterte umiddelbart den nye modellen med alle dens fortrinn og implikasjoner. Sjøl den skeptiske Rosalind Franklin var fornøyd, og hun brydde seg ikke om at hennes egne data var blitt brukt uten hennes vitende. Den 25. april 1963 ble artikkelen om DNA publisert i det berømte tidsskriftet Nature under tittelen ”Molecular structure of nucleic acids. A Structure for Deoxyribose Nucleic Acid.” Den åpner med følgende: ”Vi ønsker å foreslå en struktur for saltet av deoksyribosenukleinsyre (DNA). Denne strukturen har hittil ukjente egenskaper som er av betydelig biologisk interesse.”
Watson og Crick fikk rett! DNA har betydelig biologisk interesse. Ja, moderne biologi kunne ikke være foruten DNA. Mens Darwin ga oss den viktigste biologiske teorien, så kom Watson og Crick med den viktigste biologiske oppdagelsen.




Logg inn for å kommentere
Ikke UiO- eller Feide-bruker?
Opprett en WebID-bruker for å kommentere