- Historien om gibberellin
- Generelt om gibberellin
- Biosyntese av gibberellin i planter
- GA1 kontrollerer strekning
- Gibberellin og spiring av korn
- Gibberellin - fotoperiode og temperatur
- Andre effekter av gibberellin
Historien om gibberellin
Allerede på 1800-tallet oppdaget Japanske bønder at noen risplanter var lengre enn andre. De lange plantene var klorotiske og sterile og laget derved ikke riskorn. Sykdommen ble kalt bakanae. De første forsøkene i 1904 av Hori viste at økt strekningsvekst var forårsaket av en sopp som han trodde tilhørte slekten Fusarium. Sawada mente i 1912 at sykdommen skyldes sopphypene, men det var Kurosawa i 1926 som foreslo at det var et stoff fra soppen som ga økt strekning av plantene. Wollenweber viste i 1931 at Gibberella fujikuroi var det perfekte seksuelle stadiet og at det imperfekte aseksuelle stadiet var en art av Fusarium. I 1935 var det mulig for japanske forskere å isolere gibberellin fra filtrat fra soppkulturer av Gibberella. De laget urene krystaller som ble kalt gibberellin A og gibberellin B. Disse resultatene ble gjenoppdaget av forskere i vesten etter den 2. verdenkrig. På 1950-tallet ble strukturen til gibberellinsyre funnet av en forskergruppe ved Imperial Chemical Industries (ICI) i England og en gruppe ved USDA, Peoria i Illinois i USA. Takahashi og Tamura ved Tokyouniversitetet viste at det var tre forskjellige gibberelliner i gibberellin A: GA1, GA3 og GA2. Det viste seg at GA3 og gibberellinsyre var like. GA3 var det første kommersielle gibberellin og ble kalt gibberellinsyre. Seinere fant man gibberelliner i planter.
Idag kjenner man til flere enn 110 forskjellige gibberelliner, de fleste av dem er mellomprodukter. Gibberellinene har fått nummer i den rekkefølgen de ble oppdaget. GA1 ble først isolert fra umodne frø av bønner.
Generelt om gibberellin
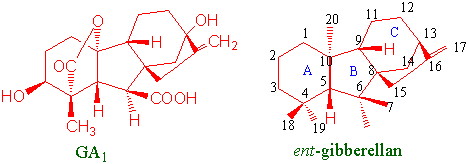
Alle gibberelliner har ent-gibberellan skjellett og flere av dem stimulerer celledeling og cellestrekning. Gibberellin tilført kunstig utenfra fremmer vekst av intakte planter.
Lokalisering av hydroksylgrupper på molekylet, oksidasjonstrinn og stereokjemi er viktig for virkningen av gibberelliner. Noen gibberelliner har 20 karbonatomer, andre har 19. Det kan være eller ikke være lakton bro mellom karbonatom 10 og karbonatom 19.
Gibberelliner finnes i hele planteriket, hos angiospermer, gymnospermer, bregner, brunalger, grønnalger, sopp og bakterier. Gibberelliner lages i unge skudd i aktiv vekst og i frø som utvikler seg. Gibberellinintermediater kan fraktes fra meristematisk vev til blad hvor sluttversjonen av gibberellin blir laget. Røtter kan lage gibberelliner.
Gibberelliner påvirker en rekke prosesser i plantene:
- - strekning av stengel og cellevekst.
- - vekst av blomsterstand og blomstring hos toårige planter som danner en rosett første året.
- - frøspiring og frøhvile.
- - partenokarpi og fruktsetting.
- - knopphvile.
- - kjønnsutrykk.
- - overgang fra ungdomsstadium til eldre.
Gibberelliner hører med til gruppen terpenoider (terpenoidbiosynteseveien) bygget opp av isopren bestående av fem karbonenheter. Forkomponenten til gibberelliner er et diterpen som inneholder 4 isoprenenheter. Det finnes en rekke veksthemmende stoffer (vekstretardanter) som hemmer biosyntesen av gibberellin og gir redusert strekning.
Veksthemmende stoffer stopper plantenes evne til å lage gibberelliner (CCC (cycocel (også kalt stråforkorter), klorocholinklorid), klormeqat, Amo 1618, phosphon D, Alar, mepiquateklorid, piperidium brom, pyrimidin forbindelser (ancymidol, A-Rest®), triazol forbindelser (paclobutrazol, Bonzil®)). Vekstretardantene har stor økonomisk betydning i hage- og landbruk. Petunia, julestjerne, liljer, Chrysanthemum og Azalea blir korte kompakte prydplanter etter behandling med stoffer som reduserer strekningen. Veksthemmende stoffer brukes også i kornåkere for å få kortere strå og mindre legde. Ønsket om redusert sprøytemiddelforbruk har gjort at planteforedlere har krysset fram kornsorter med kortere strålengde. Behandling med høyere temperatur om natten enn om dagen kan gi kortere prydplanter og redusert sprøytemiddelforbruk.
Dvergsorter av planter blir korte fordi evnen til å lage gibberelliner er redusert. Stilken hos rabarbra og stangselleri og drueklasene på vinrankene blir større og lengre etter behandling med gibberellin. Tilført gibberellin som øker stilklengden på frøløse druer gir bedre plass i klasen og derved større druer.
Jordbærplanter som sprøytes med gibberellin vil kunne gi tidligere jordbær, noe som betyr høyere pris på markedet. Nåletrær kan blomstre etter 4 - 6 år istedet for 20 hvis de blir behandlet med gibberelliner (GA4 + GA7). Dette har betydning for skogforskere som arbeider med å skaffe frø fra trær med gode egenskaper.
Siden gibberellin påvirker blomstring tror vi at biosyntese av gibberelliner må ha noe å gjøre det det hypotetiske blomstringshormonet "florigen".
Gibberelliner kan brukes i framstillingen av malt fra spirende byggkorn som skal brukes i ølproduksjon.
Umodne frø og unge blad inneholder mye gibberellin. Gibberellin fremmer spaltingen av stivelse og øker plastisiteten i celleveggen.
De vanligst brukte bioassay for gibberelliner er:
- 1) strekning av hypokotylen hos salat.
- 2) strekning av dvermutanter av ert.
- 3) strekning av nodier hos gras.
- 4) strekning av bladskjeden hos dvergris.
- 5) syntese av a-amylase i aleuronlaget hos bygg.
GA3 forekommer sjelden i planter, men kan erstatte GA1 i bioassay for gibberellin.
Biosyntese av gibberellin i planter
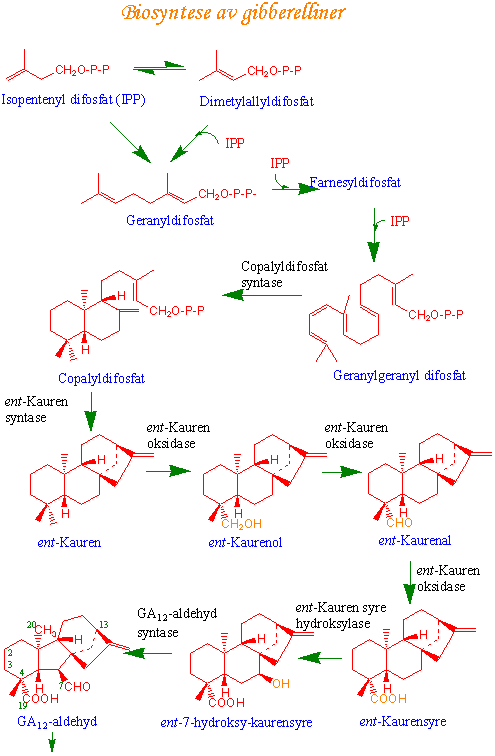
De første trinnene i syntesen av gibberelliner skjer i proplastider fra glyceraldehyd-3-fosfat, altså ikke via mevalonsyreveien. Isopren som trengs til gibberellinsyntesen lages i form av isopentenylpyrofosfat.
Mevalonsyre som kommer fra acetyl-CoA fosforylereres to ganger til mavlonsyre-5-pyrofosfat katalysert av henholdsvis mevalonsyre kinase og fosfomevalonsyre kinase. Difosfomevalonsyre dekarboksylase dekarboksylerer i en ATP-avhengig reaksjon og lager isopentenylpyrofosfat (IPP):
![]()
Isopentenylpyrofosfat isomerase katalyserer isomerisering mellom IPP og dimetylallylpyrofosfat (DMAPP). Sammenkobling av IPP og DMAPP og kondensering danner geranylpyrofosfat (C10). Geranylpyrofosfat er utgangspunkt for syntese av monoterpener, men kan reagere ytterligere med IPP og danner farnesyl pyrofosfat (C15). Farnesylpyrofosfat kan brukes til å lage sesquiterpener eller kan reagere med enda et molekyl IPP og gir til slutt geranylgeranyl pyrofosfat (C20) som kan brukes til å lage diterpener.
1. Hvordan lages ringstrukturen ?
Neste trinn er å lage en ringstruktur fra geranylgeranylpyrofosfat som ender opp i enantiomeren ent- kauren i en totrinnsreaksjon. Enzymet som katalyserer reaksjonen er en cyklase og reaksjonen skjer i proplastider i meristematisk vev. Det er dette trinnet som hemmes av vekstretardanter (AMO1618, CCC, Phosphon D - kvartære ammonium- og fosfoniumforbindelser).
2. Neste hovedtrinn er oksidasjon og syntese av GA12 aldehyd
GA12-aldehyd blir laget etter oksidasjon av metylgruppen på karbonatom 19 i ent-kauren og sammentrekning av den midterste av B-ringen fra en seksring til en femring. Metylgruppen blir oksidert til en syregruppe (-COOH) i endoplasmatisk retikulum. Oksidasjonen er katalysert av enzymet cytokrom P450 monooksygenase som kan hemmes av vekstretardanten paclobutrazol. GA12 aldehyd er forkomponent for alle andre gibberelliner.
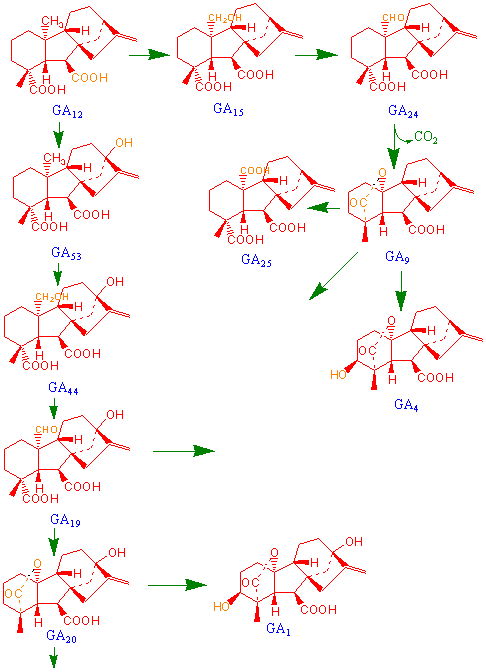
3. Andre gibberelliner lages fra GA12 aldehyd
GA12aldehyd oksideres til GA12 av enzymet monooksygenase i endoplasmatisk retikulum eller av en dioksygenase. Dioksygenase trenger 2- oksoglutarat, oksygen (O2), toverdig jern (Fe2+) og askorbat for å virke.
a) Det kan deretter skje en hydroksylering av karbonatom 13 og/eller karbonatom 3 og det dannes GA53.
b) Oksidasjon av karbonatom 20 fra metylen til aldehyd (CH2® CH2OH ® CHO (aldehyd)) katalyseres av en GA20 oksidase og GA53 går over til GA19. Deretter blir karbonatom 20 fjernet som CO2 og det dannes GA20.
GA20 hydroksyleres og gir aktivt GA1 katalysert av en 3b-hydroksylase. Deretter skjer det en 2b- hydroksylering som inaktiverer GA1 og gir GA8. GA20 kan også hydroksyleres til GA29.
Gibberelliner kan konjugeres til sukker. GA1 kan også bli uvirksom ved katabolsk inaktivering.

GA1 kontrollerer strekning
Gibberellin kan øke celledeling og cellestrekning. Det skjer ikke økt vannopptak og det skjer ikke en forsuring av veggen som ved virkning av auxin. Det ser imidlertid ut til å være en egen celleveggløsningsmekanisme som kan skyldes aktivering av enzymet xyloglukan endotransglykosylase som gjør celleveggstrukturen løsere.
Genetiske dversorter skyldes en enkelt genmutasjon slik at høyden reduseres pga. kortere internodier. Det er gjort en rekke studier av dvergmutanter av erter, mais, og ris. Hos rosetteplanter er internodiene sterkt forkortet, og når skuddet strekker seg fra rosetten skjer det en sterk økning i lengdeveksten. Hodekål består av rosetter av blad med meget korte internodier. Hvis gibberellin tilføres kunstig på bladene øker strekningsveksten (bilde).
Mendels korte og lange erteplanter
De dominante allelet Le hos ert gir lange planter. Individer som er homozygote for det recessive allelet le er dverger. Dette ble undersøkt første gang av Mendel, men det er først nå vi kan forklare hva som skjer. De høye plantene lager mer av gibberellin GA1 enn de lave, som derimot inneholder mye GA20. Dvergplantene responderer på tilført GA1 utenfra og blir da høye. Dvergene responderer derimot ikke på GA20 som er en umiddelbar forkomponent for GA1. Le allelet koder for proteinet som omdanner GA20 til GA1, en 3b-hydroksylase. Det recessive allelet le skyldes mutasjon i Le genet, men syntesen av GA1 er ikke fullstendig blokkert i dvergene (Na le). Høye erteplanter har allelene Na Le.
na er en recessiv mutant av erter hvor dannelsen av GA12 fra ent-kauren er blokkert. Planter som er homozygote for na lager ikke gibberelin og er derfor ekstremt korte, ultradverger (na na). Villtypeallelet er det dominante Na.
Det finnes også ultrahøye mutanter (la cry) av erter.
Strekning av andre arter
Dypvannsris (Oryza sativa) er delvis neddykket under vann. Internodiene strekker seg når vannivået øker og dette er styrt av gibberellin. Risrøttene har liten tilgang på oksygen og dette kan gi økt syntese av etylen.
En mutant av mais dwarf-5 er homozygot og gir svært korte planter.
Sukkerrør (Saccharum officinarum) er et eksempel på de få plantene som lagrer sukrose istedet for stivelse. Sukkerrør kan bli fra 4-6 m høye og sukrose lagres i vakuolen i parenkymceller i internodiene. Gibberellin gir økt lengde av internodiene og derved mer sukrose.
Gibberellin og spiring av korn
I aleuronlaget i korn finnes proteinlegemer og oleosomer omgitt av en halv enhetsmembran. Aleuronlaget ligger rundt endospermen med opplagsnæring. Stivelse kan spaltes til oligosakkarider inne i molekylet av enzymet a-amylase . Enzymet b-amylase kapper av oligosakkarider fra enden av stivelse og får derved flere ender å arbeide med (bilde). Det kappes av disakkaridet maltose som kan spaltes til glukose katalysert av maltase. b-amylase er tilstede i uspirt korn, mens a-amylase lages ved spiringen. Gibberellin gir økt utskillelse av enzymene b-1,3-glukanase, hydrolase, og ribonuklease fra aleuronlag og skutellum. I tillegg øker gibberellin transkripsjonen av a-amylase mRNA. a-amylase hører med til en multigenfamilie.
Gibberellin - fotoperiode og temperatur
Gibberellin deltar når knopper og frø våkner fra hvile. Økt mengde gibberellin i planten er en del av oppvåkningen. Gibberelliner kan erstatte kravet til lav temperatur og kravet til lang dag for planter som trenger dette for å blomstre. Ofte kan behandling av plantene med gibberellin gi raskere blomstring. Når planter som trenger lang dag flyttes fra kort dag til lang dag påvirker metabolismen av gibberellin. GA20 oksidase katalyserer overgangen fra GA53 til GA20 og innholdet av både GA20 og GA1 øker ved lang dag.
Gibberellin og temperatur
Lav temperatur som er en forutsetning for frøspiring hos mange planter kalles stratifisering. Lav temperatur som induserer blomstring kalles vernalisering. Gibberellin kan erstatte kuldebehandling på planter som trenger dette for stengelstrekning og blomstring. Hvis plantene ikke kuldebehandles øker innholdet av ent-kauren i skuddspissen. Etter kuldebehandling hvor plantene kommer tilbake til høy temperatur overføres ent-kauren til GA9. Det er kjent at GA9 er aktiv ifm. blomstring. Det skjer en kuldeindusert økning i ent-kaurensyre 7b-hydroksylase i skuddspissen.
Andre effekter av gibberellin
Gibberellin kan hos noen planter påvirke overgangen fra ungdomsfase (juvenilt stadium) til voksen fase. Bergflette (Hedera helix) har forskjellige bladform i ung og gammel fase. Gibberelliner regulerer begge veier.
Et juvenilt bartre kan omdannes til reproduktiv fase med behandling med GA4 + GA7.
Gibberelliner kan påvirke initieringen av blomstring og blomsterkjønn, ved abortering av enten hannlige eller hunnlige primordier. Maisplanter som får korte dager og kalde netter får økt innhold av gibberellin i kolben og gibberellin undertrykker dannelsen av hannblomster. Gibberellin kan indusere dannelsen av hannblomster i agurker. Dette kan ha betydning ved formering av F1- hybrider av agurk som vanligvis bare lager hunnblomster.