
Den gunstige effekten aske, møkk, kalk eller døde planterester har på planteveksten har vært kjent i over 2000 år. Det var Justus von Liebig (1803-1873) som samlet og systematiserte kunnskapen om grunnstoffene og deres effekt på planteveksten. Planter tar også opp grunnstoffer (elementer) som de ikke trenger og som i tillegg kan være giftige, slik at opptak av grunnstoffer er ikke noe kriterium for at plantene virkelig trenger dem i metabolismen. Vannkulturer brukes for å studere effekten av næringsstoffer på vekst og eventuelle mangelsymptomer.
{kind=link}
Nødvendige grunnstoffer
Nødvendig grunnstoff er et:
- 1) som planten trenger for å fullføre livssyklus.
- 2) hvor funksjonen til et grunnstoff ikke kan erstattes av et annet grunnstoff.
- 3) hvor grunnstoffet må delta aktivt i plantens metabolisme som en del av et kjemisk stoff, enzym, enzymreaksjon, i en struktur eller i et metabolsk trinn.
Grunnstoffene kan ordnes i et periodisk system etter antall protoner i kjernen. Det første grunnstoffet er hydrogen med ett proton i kjernen. Det neste er helium med to protoner og to nøytroner i kjernen. Hvorfor er edelgassene så lite reaktive ? Edelgassene har et stabilt antall elektroner i fylte elektronskall. De kjemiske egenskapene til grunnstoffene avhenger av antall elektroner som går rundt kjernen, slik at de enten kan avgi, motta eller dele elektroner med nærliggende atomer for å få en mest mulig stabil konfigurasjon.
Alkalimetallene (hovedgruppe I) litium, natrium, kalium, rubidium, cesium og francium er karakterisert av ekstrem kjemisk reaksjon med andre stoffer fordi de så lett avgir sitt eneste elektron i ytterste elektronskall. De gir vekk et elektron til alle stoffer som trenger det og blir enverdige i ioneform.
Jordalkalimetallene (hovedgruppe II) magnesium, kalsium, strontium, barium og radium vil gjerne gi vekk to elektroner og blir toverdige i ioneform. Plantene tar opp grunnstoffer fra jorda i form av ioner. Plantene kan også ta opp ioner gjennom bladverket.
Grunnstoffer som planten ikke trenger, men f.eks. hindrer opptak av toksiske stoffer, påvirker osmotisk trykk (natrium hos saltplanter), eller gir epidermis ekstra styrkeegenskaper (silisium) kalles nyttige grunnstoffer.
Det er stor forskjell mellom konsentrasjon av næringsstoff i jorda og det planten trenger. Jorda kan inneholde grunnstoffer i så høye konsentrasjoner at det er for mye i forhold til hva planten trenger. Derfor må opptaket være selektivt. Noen grunnstoffer tas opp aktivt, noen diskrimineres og holdes ute. Det er forskjeller mellom arter og genotyper når det gjelder ioneopptak fra jorda.
Grunnstoffene kan inndeles i:
- 1) Makronæringsstoffer som plantene trenger mye av (karbon, hydrogen, oksygen, nitrogen, fosfor, kalium, kalsium, svovel og magnesium).
- 2) Mikronæringsstoffer som plantene trenger mindre av (jern, klorid, kobber, mangan, zink, molybden, bor og nikkel).
Karbon, hydrogen og oksygen får plantene fra karbondioksid i lufta og vann i jorda. Resten av grunnstoffene tas opp som ioner (anioner og kationer) fra jorda.
En annen inndeling av mineralnæringsstoffene er i metaller og ikke-metaller:
- 1) Metaller (kalium, kalsium, magnesium, jern, mangan, zink, kobber, molybden og nikkel.
- 2) Ikke-metaller (nitrogen, svovel, fosfor, bor og klorid).
Grunnstoffer og funksjon i plantene
Grunnstoffene har som funksjon i plantene:
- 1) Inngå i alle organiske molekyler i metabolisme og strukturer (karbon, hydrogen, oksygen, nitrogen, svovel, fosfor).
- 2) Elektrontransportører i enzymer og proteiner (jern, kobber, molybden, mangan).
- 3) Regulering av metabolisme og inngå i strukturer (kalsium).
- 4) Osmotisk viktige grunnstoffer (kalium, klorid).
- 5) Koenzymer, vitaminer og andre metallenzymer (svovel, zink, magnesium).
- 6) Energimetabolisme (fosfor).
Nitrogen og svovel er en integrert del av proteiner. I de fleste enzymreaksjoner trengs en ikke-protein komponent: koenzym eller prostetisk gruppe. Transisjonsmetallene i metallproteiner virker katalytisk ved å skifte valens. Noen danner enzymsubstrat- metallkomplekser. Ioner spiller rolle ved hydratisering, stabilitet og konformasjon av enzymer. Kalium er et element som har regulatoriske rolle i enzymer ved å endre konformasjonen til proteiner. Reversibel fosforylering av enzymer via proteinkinaser og defosforylering er en viktig måte å regulere aktiviteten til mange enzymer. Flere protein kinaser trenger kalsium (Ca2+).
Noen grunnstoffer er nødvendig hos ernæringen hos mennesker, men trengs ikke eller i mindre grad hos planter. Mennesker har behov for natrium, men trengs bare hos noen saltplanter. Vi trenger litt fluorid (F-) siden fluoroapatitt reduserer nedbrytning av tennene ved karies. Selen følger svovelmetabolismen, og mennesker trenger litt selen i tilknytning til enzymet glutation peroksidase. Vi trenger jodid for tyroidhormonet i skjoldbruskkjertelen.
Mineralnæring historie
Flom i Nilen ga avsetning av leire og mineralrikt slam som kom fra høylandet i Etiopia og Sudan, og ga grunnlag for landbruk og sivilisasjonen i det gamle Egypt, på samme måte som for de andre elvekulturene. Det ble bygget avanserte vannkanalsystemer (akvadukter), også av aztekerne i Mexico med sine flytende hager. Fra gammelt av visste man at nedhogging og brenning av skog (svedjebruk, svijordbruk) kunne gi næringsrik jord for dyrking av for eksempel svedjerug. Ved å la jorda deretter ligge brakk noen vekstsesonger vokste det opp ugras som ga tilførsel av organisk materiale (grønngjødsling), med ny mulighet for dyrking av rug. John Woodward (1665-1778) oppdaget i 1699 at vann fra Themsen ga bedre vekst av planter sammenlignet med regnvann.
Den engelske kjemikere Joseph Henry Gilbert (1817-1901) og John Bennet Lawes (1814-1900) startet i 1837 eksperimenter med effekten av avføring fra husdyr på plantevekst. Lawes Estate ble til Rothamsted Research Station, The Rothamsted Experiments (1895). Man fant at bein og fosfatstein ga bedre vekst hvis det først ble behandlet med svovelsyre, noe som ga superfosfat (P2O5). Biologen og statistikeren RA Fisher bidro seinere i betydelig grad til design av landbrukseksperimentene, og statistiske metoder for analyse av forsøksresultatene.
Alexander von Humbolts reiser ga på begynnelsen av 1800-tallet oppdagelsen av guano på øyer utenfor kysten av Peru. Guano ble også funnet på øyer i Karibien, utenfor kysten av Namibia, Oman og Patagonia. I områder med lite nedbør ga avføring fra fuglekolonier tykke lag med guano. Det ble også brukt guano fra flaggermus funnet i fjellhuler. Guano inneholder ammoniumnitrat, urinsyre, fosfat og andre mineraler. Guano innholder mange småkryp og mikroorganismer, blant annet soppen Histoplasma capsulatum som inneholder sporer som gir lungesykdommen histoplasmose. Histoplasma har to morfer, en ved lav ytre temperatur og en ved kroppstemperatur hvor de kan leve i fagosomer. Guano ble fraktet til Europa, og ga grunnlaget for det europeiske landbruket med tilhørende befolkningsøkning. Amerika sikret seg i 1856 rettigheter til guano via Guano Island Act. Den tyske kjemikeren Justus von Liebig (1803-1873), samt den franske kjemikeren Jean-Baptiste Boussingault (1801-1887) som fikk bygget en forsøksstasjon for landbruk nær Alsace, bidro til utvikling av landbruktsforskningen.
Liebig viste at nitrogen, fosfor og svovel er viktig for plantevekst, og at veksten er begrenset av det næringsstoffet det er minst av, bl.a. omtalt i boka Die Organische Chemie in ihre Anwendung auf Agrikultur und Physiologie (1841) (Organisk kjemi og dens anvendelser innen landbruk og fysiologi). Aske fra brent plantemateriale ble kalt pottaske og inneholder kaliumkarbonat . På 1860-tallet oppdaget man i Tyskland avsetninger av pottaske (K2O), seinere også andre steder som ble brukt i gjødsel.
Guanoforekomstene ble tømt, men ble erstattet på 1870-tallet av chilesalpeter fra Atacamaørkenen. I kjølvannet av nitrogenforekomstene fulgte Guanokrigen og Salpeterkrigen. Chilesalpeter ble erstattet av Birkeland og Eides Norgessalpeter. Den tyske kjemikeren Fritz Haber (1868-1934) viste at med hjelp av en jernkatalysator var det mulig å lage ammoniakk (NH3) fra hydrogen (H2) og nitrogen (N2) . Haber-Bosch-prosessen i samarbeid med Carl Bosch (1874-1940) ved BASF ble prosessen videreutviklet ved bruk av høyt trykk og temperatur .
Hydroponikk, dyrking av planter uten jord i vannkultur ga videre identifisering av de andre nødvendige næringsstoffene. Den tyske plantefysiologen Fardinand Gustav Julius von Sachs (1832-1897) laget oppskrift på en næringsløsning brukt i vannkultur. Dyrkningsteknikken hydroponikk ble perfeksjonert av Wilhelm Knop (1817-1891) som plasserte frøplanter festet med en bomullspropp i hull i korker som fløt på Knops næringsløsning i dyrkningskar. Knop kunne vise at planter er ikke avhengig av jord for å kunne vokse, de trenger bare de uorganiske grunnstoffene i form av mineraler i jorda. William F Gericke innførte begrepet hydroponikk i boka The complete guide to soilless gardening (1940).
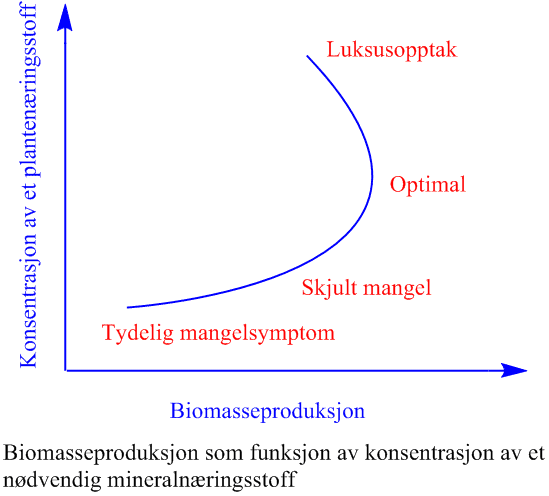
Innholdet av makronæringsstoffene nitrogen (N), fosfor (P), kalium (K), svovel (S), magnesium (Mg) og kalsium (Ca) i plantemateriale varierer fra 1 -20 mg per gram friskvekt, mens mikronæringsstoffene er i mengder mikrogram per gram friskvekt.
pH og ioneopptak
Reaksjon mellom protoner, hydroksylioner, bikarbonat og andre kationer vil påvirke ioneopptaket. pH i jorda er sjelden over pH 7.5 slik at protonkonsentrasjonen vil ha stor betydning. Når den ytre protonkonsentrasjonen øker vil effluksen av protoner reduseres f.eks. kan et membranpotensial på -150 mV ved pH 6 bli redusert til -100 mV ved pH 4. Det er forskjell på uladete molekyler og ladete anioner og kationer når det gjelder å kunne komme gjennom biomembraner. Det betyr f.eks. at ioner som H2PO4- tas lettere opp enn HPO42- slik at dette opptaket blir pH avhengig. I sur jord bedres tilgangen på jern og mangan betydelig sammenlignet med mer basisk jord.
Det finnes to systemer for regulering av pH i plantene:
- 1) Protonutbytting over membraner ved ATPase, ATP syntase og antiporttransport.
- 2) Biokjemiske pH regulering via produksjon og bruk av protoner ved dannelse og fjerning av karboksylsyrer.
Det vesentlige av det selektive opptaket skjer i plasmamembranen. Den andre barrieren er tonoplasten. Protonpumper i membranen regulerer pH i cytoplasma, men gir samtidig en elektrokjemisk gradient som kan brukes til å transportere andre stoffer: nøytrale, anioner og kationer. I rothår er det beregnet at H+ATPase bruker opptil 50 % av cellens ATP. H+ATPase aktiveres også av K+. Kationer kan transporteres i en nedoverbakkeprosess i en uniport av bærere (permeaser) eller ved kotransport. For aniontransport kan man ha en proton-anion kotransport (symport) som bruker den elektrokjemiske gradienten som drivkraft. Fosfat kan tas opp av en OH--fosfat mottransport (antiport). Protonpumpene er fosfohydrolaser som enten bruker pyrofosfat eller ATP som energikilde. Disse trenger Mg2+, noe som indikerer at MgATP eller MgPP er substrater. MgPP-ase trenger også K+. Protonpumpen i tonoplast sørger for høy pH i cytoplasma og lav pH i vakuolen, og transporterer kationer f.eks K+ for å opprettholde det osmotiske trykket. Tonoplasten har ATPase og PPase, plasmamembranen har ATPase.
Stort opptak av kationer følges av økt pH i cytoplasma og induserer syntese av organiske syrer. Høyt opptak av anioner gir lavere pH i cytoplasma (proton-anion-kotransport). For å opprettholde høy pH i cytoplasma trengs økt dekarboksylering av organiske syrer fra f.eks. vakuolen. Likevekten mellom karboksylering (CO2-fiksering) og dekarboksylering reguleres av pH avhengigheten for enzymene fosfoenolpyruvat karboksylase og malatenzym. En økning i pH aktiverer fosfoenolpyruvat karboksylase og ved hjelp av malat dehydrogenase lages malat som kan lagres i vakuolen hvor den virker som motion for kationer. Vi har funnet høye konsentrasjoner av malat i stengelen hos unge kimplanter av gran. Hvis pH i cytoplasma minsker blir malatenzym aktivert, noe som leder til dekarboksylering av malat (aplesyre) og produksjon av CO2. I planter som lager mye oksalsyre kan denne være med i pH reguleringen. Kalsium-oksalatkrystaller er vanlig i nålene hos bartrær, i celleveggene i mesofyll og floem og i ytterveggen av epidermis. Oksalsyre kan brukes til å fjerne overskudd av kalsium eller som antibeitestoff. Oksalsyre kan også brukes til å lage hydrogenperoksid som brukes til bekjempelse av sykdomsfremmkallende organismer. Opptaket av ammonium og nitrat er det som har størst betydning for pH. Surjordsplanter som foretrekker ammonium som nitrogenkilde har et høyt kation/anion forhold sammenlignet med planter som tar opp nitrat.
Ammoniumassimilasjon produserer protoner: 3 NH4+ assimilert gir 3 RNH2 + 4 H+. Protonene kan skilles ut fra rota i bytte med ammonium hvis assimilasjonen skjer i rota. Assimilasjon av nitrat gir tilsvarende mengder hydroksylioner (OH-). 3 NO3- gir 3 RNH2 + 2 OH-. pH reguleringen blir enklest hvis plantene har tilgang til både nitrat og ammonium.
Transportveien inn i rot og celler
Hovedbarrieren for ionefluksen i apoplasten i rota er endodermis med hydrofobe Casparyske bånd. Suberin i celleveggen i endodermis er en effektiv barriere for videre transport i apoplasten. I de fleste planter finnes det et cellelag i rotbarken (cortex) i form av en hypodermis eller eksodermis som har suberin i veggene, og som fungerer som en barriere på lik linje med endodermis. Etter radial transport inn til stelen vil ioner og organiske stoffer (aminosyrer, organiske syrer) fraktes inn i xylemet langs en elektrokjemisk gradient som er koblet til respiratoriske prosesser. Det er ting ved denne innlastingen i xylemet som fremdeles er ukjent. I motsetning til xylemet går transporten i floemet begge veier, opp og ned i planten. Når mineralnæringsstoffene har kommet inn i planten kan flere av dem refordeles i planten via floemet f.eks. nitrogen, fosfor og kalium, men jern og kalsium er eksempler på stoffer som ikke kan retranslokeres på denne måten. Det kan skje reabsorbsjon av mineralnærings langs transsportveien oppover i planten over til parenkymceller, men mesteparten fraktes til bladene.
Bevegelsen av lavmolekylære stoffer, ioner, organiske syrer, aminosyrer og sukker fra den ytre løsning og inn i celleveggene i rotcellene, - det frie diffusjonrom, skjer som en passiv ikkemetabolsk prosess. Celleveggen kan imidlertid reagere med stoffene og fremme eller hindre videre bevegelse innover i planten. I celleveggens nettverk er det interfibrillære og intermicellære rom i størrelsesorden fra 3- 5 nm, sammenlignet med hydrert kalium (K+) som har diameter 0.66 nm og kalsium (Ca2+) med diameter 0,82 nm. Høymolekylære stoffer som metallchelater, fulvinsyrer, og virus kommer ikke gjennom porene. I dette nettverket hvor det er mye pektin (polygalakturonsyre), spesielt i midtlamellen, blir det et kontinuerlig apoplasma hvor karboksylsyregrupper (RCOO-) virker som kationbyttere. Siden disse ladningene ikke representerer noe fritt diffusjonsrom kalles de tilsynelatende fritt diffusjonsrom. Dette består av et "waterfreespace" som er tilgjengelig for ioner og ladete og uladete molekyler, og "Donnan fri space" hvor det skjer anion- og kationbytte. Plantene varierer i evne til kationbyttekapasitet, det vil si hvor mange fast bundete anioner (RCOO-) det finnes i celleveggene. Kationbytterkapasiteten er noe høyere for tofrøbladete enn for enfrøbladete planter. Kationbytterkapasiteten minsker når den ytre pH i jorda blir surere. I membranene vil fosfo-, glyko- og sulfolipider, og proteiner inneholde elektriske ladninger som vil tiltrekke seg ioner med motsatt ladning. Ionekanaler er spesielle proteiner i membranen som gir stor innfluks av ioner som er 3-10 x raskere enn for bærerproteiner (permeaser). Makromolekyler kan tas opp i form av endocytose (pinocytose). Transportproteinene i cellemembranene har en opptakskinetikk som ligner enzymkinetikk og man bruker ofte de tilsvarende. Siden transportproteinene ikke endrer substratet kan de ikke kalles enzymer. Spesielt for natrium (Na+) og kalsium (Ca2+) er det stor forskjell mellom beregnet konsentrasjon i cellen ut fra Nernst-ligningen og det som måles, noe som indikerer at begge disse blir aktivt transportert ut av cellene.
Konkurranse om opptak
Den metabolske aktiviteten i røttene har betydning for opptaket slik at oksygentilgang og mengde karbohydrater i røttene blir viktig. Det samme gjelder rottemperatur og konkurransen mellom grunnstoffer som har samme valens og ionediameter. Det er konkurranse mellom opptak av alkalikationene kalium (K+), rubidium (Rb+), og cesium (Cs+). Bindingen av cesium er relativt lav. Det betyr at kalium kan utkonkurrene cesium, men ikke omvendt. Det har betydning for opptak av radioaktivt 137Cs inn i planter hvor cesium utkonkurreres fra ioneopptak på dyrket mark med god kaliumgjødsling, men ikke i næringsfattig utmark og i fjellområder. Derfor vil sauer på utmarksbeite inneholde radioaktivt cesium i minst 50 år etter Tsjernobylulykken i 1986. Neste utslipp var ved kjernekraftverket Fukushima i Japan etter en tsunami.
Ammonium (NH4+) kan hemme opptaket av kalium (K+), men ikke omvendt. Både kalium (K+) og kalsium (Ca2+) hemmer opptaket av magnesium (Mg2+). Det vil si sterk gjødsling med kalium og kalking kan gi magnesiummangel. Mangan (Mn2+) kan hemme opptaket av Mg2+. Det er også konkurranse i opptak mellom molybdat (MoO42-), sulfat (SO42-), selenat (SeO42-),og mellom fosfat (HPO42-) og arsenat (H2AsO42-). Økt mengde sulfat hemmer opptaket av molybdat og selenat. Arsenat og fosfat tas opp av samme opptakssystem. Dette er av betydning hvis jorda inneholder mye molybden, arsen eller selen. Arsen kan komme fra trykkimpregnert treverk. I noen geografiske områder kan jorda inneholde mye selen. Selen inkorporeres i stedet for svovel inn organiske forbindelser f.eks. aminosyrene selenomethionin, selenocystein.
Det er også konkurranse mellom klorid og nitrat. Klorid innholdet kan reduseres i planter ved å tilføre mye nitrat. Omvendt kan man senke nitratinnholdet i planter som spinat ved å tilføre mye klorid. Ammonium vil ofte undertrykke opptaket av nitrat, men ikke omvendt. Opptak av ammonium vil gi redusert membranpotensial og vil derfor hemme nitratinfluks Det vil være en konkurranse mellom H+ og aluminiumkomplekser og opptaket av andre elementer som f.eks. magnesium. ZnEDTA kan gi mindre zinkopptak enn hvis man har Zn2+ . Kalsium (Ca2+) tilførsel vil hindre at kalium (K+)- opptaket reduseres ved lav pH. Hos natriumfobe planter som bønner vil natrium (Na+) forbli lokalisert til røttene, mens natriumfile planter som sukkerbete vil transportere natrium opp i bladene.
Retranslokasjon av grunnstoffer via floemet
Grunnstoffene fra jorda tas opp i plantene via transpirasjonsstrømmen i vedvevet (xyleme. I ledningsstrengene er xylem (vedvev) og floem (silvev) plassert inntil hverandre. Nær kontakt mellom xylem og floem gjør at grunnstoffer kan forflyttes mellom disse vevene. Flere av grunnstoffene fra skuddet kan retranslokeres tilbake til røttene eller til de yngste delene av planten. En del av de retranslokerte grunnstoffene blir ført tilbake igjen til xylemet i røttene, og på den måten resirkulert i planten. Høyere alder på bladene resulterer i større eksport enn import av grunnstoffer, og det skjer en refordeling og remobilisering av grunnstoffene internt i plantene. For rasktvoksende åkervekster blir remobilisering fra eldre blad til de yngre avgjørende. Hvis næringsmangelsymptomer viser seg først på de yngste bladene tyder dette på lite effektiv remobilisering. Remobilisering av grunnstoffer blir spesielt viktig ved reproduktiv vekst når det skal lages frø, frukt og lagringsorganer. I reproduktiv fase synker innholdet av grunnstoffer i de vegetative delene. Korn har f.eks. høyt innhold av nitrogen og fosfor, og lave konsentrasjoner av kalium, magnesium og kalsium, mens saftige frukter og lagringsorganer ofte har et høyt innhold av kalium, men relativt lite nitrogen og fosfor. Remobiliseringen kan i noen tilfeller være så aktiv at det virker som et selvdestruerende system. Remobilisering av grunnstoffer er vanlig før bladfall om høsten hvor nitrogen, kalium, fosfor og zink retranslokeres aktivt. I natrofobe planter er retranslokalasjon en måte å holde lave konsentrasjoner av natrium i bladene. I planter hvor nitratreduksjonen skjer i skuddet, må ammonium kunne bli transportert tilbake til røttene. Sirkulasjon av kalium kan være med å skape ladningsbalanse for planter som bruker nitrat som viktigste nitrogenkilde, hvor kalium er et viktig motion for nitrattransporten. Ved frøspiring remobiliseres grunnstoffer fra opplagsnæringen, spesielt kalium, magnesium og kalsium bundet til fytinsyre. Frigivelsen av fosfat følger aktiviteten til enzymet fytase.