Fosfat finnes i pH-likevekter i form fra fosforsyre (H3PO4) til fosfat (PO43-) ifølge et Bjerrumdiagram
H3PO4 ↔ H2PO4- ↔ HPO42- ↔ PO43-
Ved høy pH i jordsmonnet kan fosfat finnes som kalsiumsaltet hydroksyapatitt og
i sur jord som aluminiumfosfat eller jernsulfat, og fosfat tes lettest opp ved pH 6-7
Eksempler på fosfatplanter som tives i fosfatrikt jordsmonn er svaleurt (Chenopodium majus, daunesle (Lamium album) og stolt-henrik (Chenopodium bonus-henricus)
Fosfor kan kontrollere immobilisering av karbon og nitrogen i marine miljøer fordi Redfieldforholdet C:N:P er 106:16:1. C:N.P støkiometri har en viktig funksjon i biogeokjemiske sykluser. Fosfor kan være en begrensende faktor for vekst og plantene har utviklet strategier for å effektivisere opptaket av fosfor. Det skjedde en endring i fosfatkonsentrasjonen i havet da plantene koloniserte land. Bakterier, sopp, alger og protozooer kan lagre fosfor som polyfosfat, også kalt volutin. Fosfat lagres i frø som fytat, som blir spatet av fytase ved frøspring.
Fosfor kan binde seg til et oksygen- (O) eller karbonatom (C). Energimetabolismen består av fosforylerte organiske forbindelser. Oksygenet i fosfat kan danne et fosfoanhydrid, et difosfat (e.g. pyrofosfat og ADP) eller hvis tre fosfat, et trifosfat f.eks. ATP og GTP. I et bisfosfat er fosfor koblet til forskjellige karbonatomer f.eks. ribulose-1,5-bisfosfat i assimilasjon av CO2 i Calvinsyklus i fotosyntesen. Fosfor kan også bindes til et organisk molekyl i en fosfodiesterbinding, slik som i DNA og RNA.
| Fosforylert forbindelse | \(\Delta G^ {0'}\)kJ mol-1 |
|---|---|
| ATP + H2O→ADP + Pi (uorganisk fosfat) | -30.5 |
| Fosfodiester (PPi (pyrofosfat) → Pi | -25.1 |
| AMP + H2O→ adenosin + Pi | -13.8 |
| Glukose-6-fosfat → glukose + Pi | -13.8 |
| Glukose-1-fosfat → glukose + Pi | -20.9 |
| Fosfoenolpyruvat | -54.4 |
| Acetyl-fosfat | -43.9 |
| 1,2-bisfosfoglycerat | -49.4 |
| Kreatinfosfat | -37.7 |
Endring i Gibbs fri energi \(\Delta G^{0'}\)ved hydrolyse av fosforylerte forbindelser med standardbetingelser og pH 7.
Eksempler på andre fosforylerte forbindelser i metabolismen er i glykolysen fruktose-6-fosfat, fruktose-1,6-bisfosfat, glyceraldehyd-3-fosfat også som isomer med hihydroksyacetonfosfat, 1,3-bisfosfoglycerat, 3-fosfoglycerat, 2-fosfoglycerat og fosfoenolpyruvat. I glykolysen inneholder både 1,3-bisfosfoglycersyre og fosfoenolpyruvat nok energi til å lage ATP, en substratnivpfosforylering, katalysert av henholdsvis enzymene fosfoglycerat kinase og pyruvat kinase. I sitronsyresyklus (Krebs-syklys, trikarboksylsyresyklus) brukes den fri energien i succinyl-CoA til å lage ATP, katalysert av succinyl-CoA syntetase hvor det andre produktet blir succinat.
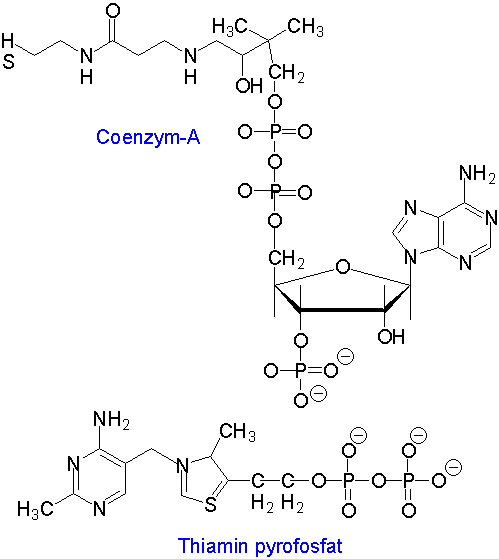
Koenzym A og thiamin pyrofosfat er eksempler på fosforylerte koenzymer.
ATP blir laget kjemiosmotisk ved fotosyntetisk fosforylering i kloroplastene, respiratorisk fosforylering i respirasjonskjeden i mitokondriene, katalysert av ATP-syntase. H+-ATPase bruker den fri energien ved hydrolyse av ATP til å frakte protoner over en membran og senker membranpotensialet i kompartmentet.
Orbitalstruktur for fosfor: 1s2 2s2 2p6 3s2 3p3 = [Ne] 3s2 3p3
Fosforassimilasjon
Fosfat behøver ikke reduseres før assimilering inn i organiske forbindelser, i motsetning til nitrat og sulfat. Fosfat tas opp i røttene med HPO42- H+ symporter. Etter å ha blitt tatt opp i plantene som H2PO4- forblir fosfat som uorganisk fosfat (Pi, i="inorganic") eller fosfat kan bli forestret til en hydroksylgruppe (CO-P) som en fosfatester eller energirik polyfosfat f.eks. pyrofosfat (PPi). I mange reaksjoner er fosfat et substrat eller produkt. Vakuolen er lagringssted for ikkemetabolsk fosfat. Fosfor inngår en rekke steder i metabolismen.
Fosfor inngår i nukleinsyrene DNA og RNA. Det er fosfor som gir de sure egenskapene til nukleinsyrene. Fosfatgruppen vil ha negativ ladning ved nøytral pH. Et nukleotid består av en base, sukker og fosfat, og fosfat er bundet til sukker i en esterbinding.
Fosfor inngår i energimetabolismen som sukkerfosfatintermediater i respirasjon og fotosyntese. Hydrolyse av fosfatesterbindingen til fosfat frigir store mengder energi fra høyenergimolekyler. Hydrolyse av ATP katalysert av enzymet ATPase gir ca. 30 kJ mol-1. En fosforylgruppe i ATP kan overføres til andre molekyler. Andre energirike fosfatestere er fosfoenolpyruvat (-62 kJ mol-1), pyrofosfat (-28 kJ mol-1) og glukose-6-fosfat (-14 kJ mol-1). ATP lages ved substratfosforylering i respirasjonen eller ved fotofosforylering i fotosyntesen katalysert av enzymet ATPsyntase.
Fosfor inngår i fosfolipider i biomembraner, Fosfodiestere i form av fosfolipider er vanlig i alle biomembraner f.eks. som fosfatidylkolin (lecithin). Fosfolipidene har en hydrofob hale dannet av to fettsyrer som vender inn i membranen og et hydrofilt hode som vender ut mot cytosol eller stroma.
Fosfor deltar i signaloverføringsveier i metabolismen ved fosforylering og defosforylering av proteiner katalysert av henholdsvis protein kinaser og defosforylaser. GTP og GDP inngår i G-proteiner. Plantene inneholder hundrevis av forskjellige protein kinaser, og opptil 4% av genomet kan være involvert i syntesen. Andre signalmetabolitter er syklisk AMP (cAMP) og syklisk GMP (cGMP), hvorav den sistnevnte er viktigst i planter.
Transporten av triosefosfater ut av kloroplasten skjer via en fosfattranslokator i bytte mot fosfat. Fosfattranslokatoren sitter i den indre kloroplastmembranen. Syntesen av stivelse hemmes av høye konsentrasjoner av fosfat. Enzymet for syntese av stivelse i kloroplasten Enzymet ADP-glukose pyrofosforylase som lager stivelse i kloroplastene hemmes av uorganisk fosfat og aktiveres av triosefosfater.
Stivelse fosforylase er et reversibelt enzym i plastider:
α-1,4-glukann (stivelse) + Pi → glukose-1-fosfat + α-1,4-glukan(n-1)
Fytat
Fytat er et lager med fosfat og andre grunnstoffer. Fosfat lagres som fytat (fytin) i pollen, frø og korn. Fytat er et kalsium-magnesiumsalt av fytinsyre. Fytinsyre lages fra myo-inositol ved at hydroksylgruppene er forestret med fosfat. Fytinsyre (inositol heksafosforsyre) har også høy affinitet til andre grunnstoffer som jern og zink. Fytat spaltes av enzymet fytase. Syntesen av fytat i frø regulerer også stivelsesyntese ved å senke fosfatkonsentrasjonen via reaksjonen med fytat.
Fosfat
Fosfat (PO43-) med tre negative ladninger er et atom fosfor omgitt av fire oksygen i et tetraeder.
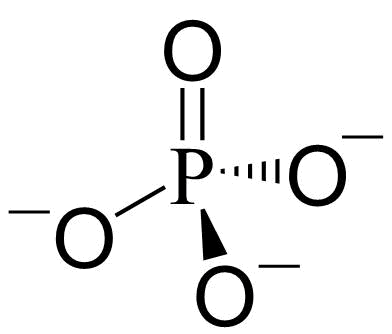
Fosfat er saltet av fosforsyre. Fosfat (ortofosfat, PO43-) er konjugert base til hydrogenfosfat (HPO42-), som igjen er konjugert base til dihydrogenfosfat (H2PO42-). Dihydrogenfosfat (H2PO42-) er konjugert base til fosforsyre (H3PO4). Ved fysiologisk pH er det en likevekt mellom HPO42- og H2PO4-. Ved 25oC er likevektene (Ka i mol l-1) med de tilsvarende pKa-verdiene (log10 til likevektskonstanten)
H3PO4 ↔ H+ + H2PO4- (Ka1= 7.5·10-3, pKa1 =2.12)
H2PO4- ↔ H+ + HPO42- (Ka2 = 6.2·10-8, pKa2= 7.21)
HPO42- ↔ H+ + PO43- (Ka3 = 2.14·10-13, pKa3 = 12.67)
Ved pH i løsningen lik pKa-verdien er det like store mengder av de to formene av fosfat i likevekten.pKa
| Likevekt | pKa |
|---|---|
| H3PO4 ↔ H2PO4- + H+ | 2.15 |
| H2PO4- ↔ HPO42- + H+ | 7.20 |
| HPO42- ↔ PO43- + H+ | 12.37 |
Fosfat forkommer også som pyrofosfat (difosfat, P-O-P, PPi, i - inorganic, gr. pyro - ild) og polyfosfat ([PO4]n),
I biologisk materiale finnes fosfat som fritt fosfat eller bundet i organiske forbindelser. Fosfat inngår i organiske forbindelser som en ester. Fosforylering og defosforylering av proteiner er viktige i regulering av metabolismen. Kan øke tilgangen på fosfor ved å skille ut fosfatase og fosfodiesterase som bryter ned organiske fosforforbindelser. Fosfodiesterbindinger finner man i nukleinsyrer og fosfolipider, mens det er fosfomonoesterbindinger i sukkerfosfater og ATP. Røttene skiller i citrat i rhizosfæren, og piscidinsyre fra dueert (Cajanus cajan) er eksempel på en syre som løser opp jernfosfat. Fosfor finnes lagret som fytat (inositol heksafosfat) i frø som binder kalsium (Ca2+), magnesium (Mg2+), jern (Fe3+) og zink (Zn2+), og blir nedbrutt av enzymet fytase. Mikroorganismer kan også inneholde fytase. Fosfat kan gi eutrofiering i ferskvann jfr. Mjøsaksjonen. Naturlig vegetasjon har effektivt opptak av fosfor, men det har ikke landbruksvekster, noe som gir avrenning av fosfor til ferskvann. Fosformengden i landbruksjord gjødslet med kunstgjødsel øker mer enn det plantene kan ta opp. Fosfor er lite løselig og renner ikke ned i grunnvannet som nitrat, men kommer ut i vann og kystområder via erosjon av jordkolloider fra landbruksjord, blant annet fra drenering. Fosfor blir også tilført vann og vassdrag fra kloakk, husdyrgjødsel og vaskemidler.
Det er interaksjon mellom sulfat (sulfathomeostase) og fosfat (fosforhomeostase) via den MYB-relaterte transkripsjonsfaktoren PHR1 (fosfatrespons 1). Ved fosfatmangel stopper strekningen av primærroten, mens det blir økt siderotdannelse, samt gir økt tetthet og lengde av rothårene, som gir utseende som en flaskekost. Ved fosfatmangel kan fosfolipider i membranen bli erstattet med sulfolipider.
Fosfat er viktig i energiomsetningen i cellene via ATP, GTP, fosforylert glukose og fruktose, og fosforylerte triosefosfater. I membraner inngår fosfor i fosfolipider. Fosfat er en viktig buffer i cellen.Fosfat kan finnes som en polymer i form av pyrofosfat (PPi, P2O74-) eller trifosfat (PPPi, P3O105-). Hydrolyse av pyrofosfat er katalysert av en pyrofosfatase hvor det blir frigitt energi:
P2O74- + H2O ↔ 2HPO42-
Fosfatgjødsel
Fosfat tas opp av røttene via H+-fosfat symport via en superfamilie MFS transportører som ikke har et ATP-bindende sete slik som ABC-transportørene. PHT1 er en opptakstransportør i plasmamembranen med høy affinitet til fosfat, tilsvarende SULTR1 for sulfat (SO42-). Dihydrogenfosfat (H2PO4-) er lettere løselig enn hydrogenfosfat (HPO42-) og ortofosfat (PO43+) som letter gir utfellinger med kalsium (Ca2+), aluminium (Al3+) og jern (Fe2+). Fosfatkonsentrasjonen i jord er fra 0.1 til 10 µM. Kalkrik jord inneholder mye kalsiumfosfat, mens i sur jord finnes fosfor bundet til jern-aluminiumhydroksider og oksider. Jord i tropiske strøk som andisol, ultisol og oksisol inneholder lite fosfat og det blir konkurranse om tilgangen på fosfor.Fosfat er en begrenset ressurs som utvinnes fra fosfatavsetninger i form av apatitt. Imidlertid kan slike fosfatforekomster også inneholde tungmetaller som kadmium (Cd), bly (Pb), nikkel (Ni), kobber (Cu), krom (Cr) og uran (U). Kontaminert gjødsel kan gi utlekking til grunnvann. Fosfat sammen med nitrogen (N) er to grunnstoffer som begrenser plantevekst i landbruk og terrestre økosystemer. Nitrogen begrenser ofte veksten i sjøvann, mens fosfat begrenser veksten i ferskvann. I terrestre økosystemer bidrar utskillelse av fosfatase og mykorrhiza til økt opptak av fosfat i røttene.
Avføring fra fugl er rik på fosfor og nitrogen, og guano fra Chincha-øyene utenfor Peru var på 1800-tallet en viktig gjødsel for landbruket i Europa, siden den inneholder både nitrogen, fosfor og organisk stoff. Tørt klima og den kalde Humboldtstrømmen gjorde at det ble dannet tykke lag med guano. På 1900-tallet ble guano fra Naura i Stillehavet viktig. Justus for Liebigs undersøkelse av plantenes opptak av mineralnæring fra jorda ga etter hvert et grunnlag for mineralgjødsel som erstatning for guano fra fugl og flaggermus. Organisk fosfat i kompost er en annen viktig fosforkilde.
Planter har et høyt C:P-forhold og drøvtyggere har mikroorganismer i vomma som bryter ned cellulose, men tar også effektivt opp fosfor. Enmagedyr som fugler, hester og mennesker har et lite effektivt opptak av fosfor og får derfor høyt innhold av fosfor i avføringen. Når verdens fosfatforekomster minsker er det flere som mener at man må ta i bruk avføring fra mennesker som kilde til fosfor i landbruket.
Apatitt fra gruver ble behandlet med svovelsyre og ga superfosfat
Steinforsfat (Ca3(PO4)2 )+ svovelsyre (H2SO4) → superfosfat (Ca(H2PO4)2) + P2O5
Fosfatfjell behandlet med fosforsyre (H3PO4) ga trippel superfosfat, men etter hvert overtok ammoniumfosfat (NH4)3PO4).
I Oddaprosessen anvendes salpetersyre istedet for svovelsyre:
Ca3(PO4)2 + 6HNO3 + 12H2O → 3Ca(NO3)2 + 2H3PO4 + 12H2O
Apaptitt Ca10(PO4)6(OH, Cl, F)2 finnes i form av hydroksyapaptitt, kloroapatitt eller fluoroapatitt, det som vi gjerne vil ha i tennene, men fluorid i større konsentrasjoner er giftig for planter og må fjernes som flussyre (HF). HF reagerer med slikikater. Hydroksylgruppene kan også bli erstattet med karbonat.
Fluoroapatitt behandlet med svovelsyre gir reaksjonen:
CaF2·3Ca3(PO4)2 + 7 H2SO4 + 14 H2O → 3 Ca(H2PO4)2 + 7 CaSO4·2H2O + 2 HF
Eller som trippel superfosfat:
CaF2·3Ca3(PO4)2 + 14 H3PO4 → 10 Ca(H2PO4)2 + 2 HF
Fosforsyre kan utvinnes fra Ca(H2PO4)2.
Gips (CaSO4·2H2O) produsert i produksjon av fosfatgjødsel gir et miljøproblem, og kan også inneholde noe radioaktivitet fra den radioaktive serien fra uran-238. Flere millioner tonn med fosfogips som man ikke vet hvor man skal gjøre av gir problemer med å selge gjødselproduksjon som bærekraftig.
238U → 234Th → 234Pa → 234U → 230Th → 226Pa → 222Rn
Søve gruve ved Fensfeltet ved Ulefoss i Telemark inneholder søvitt, en kalkstein som inneholder nioboksid, apatitt, magnetitt, svovelkis, men også mindre mengder radioaktivt uran og thorium. Under andre verdenskrig hadde Norsk Hydro planer om å utnytte apatitten som fosforkilde i gjødselproduksjon, men etter krigen det var innholdet av grunnstoffet og metallet niob som ble mest interessant og utvunnet, inntil gruven ble nedlagt i 1963. I slagghaugene er det imidlertid forhøyet konsentrasjoner av radioaktivt materiale som nå må dekommisjoneres.
Før eller siden blir verdens lagre av apatitt brukt opp, og det meste kommer i dag fra Vest-Sahara, S-Afrika, USA, Kina og Russland.
Fosfor i jord og mangelsymptomer i planter
Fosfor er sterkt bundet i jorda, til jern (Fe3+) og aluminium (Al3+) i sur jord og til kalsium (Ca2+) i mer basisk jord. I moldjord finnes en stor del av fosforreservene i organisk form. I forvitret sur tropisk jord som oksisol (oksid henspiller på oksidmineraler som bauxitt) , ultisol (rød leirjord) og andisol (vulkansk aske) er det lite fosfor.
Ved dårlig tilgang på fosfat kan planterøttene skille ut enzymet fosfatase til jorda hvor enzymet hydrolyserer organiske fosforforbindelser. Røttene skiller også ut organiske syrer og protoner, og rothårene blir lenger ved fosfatmangel. Mykorrhiza kan lette opptaket av fosfor i røttene. Fosfor fraktes lett fra eldre til yngre deler av planten slik at det er de eldste bladene som viser mangelsymptomer hvis det blir for lite fosfor. Ved fosformangel blir de yngste bladene mørkegrønne, de eldste visner og det kan lages anthycyaniner på samme måte som ved nitrogenmangel.Plantene må konkurrere med mikroorganismene i jorda om tilgangen på fosfat.
Ved mangel på fosfor hemmes veksten av skuddet, men rotveksten hemmes ikke i samme grad, slik at det blir økt rotbiomasse på bekostning av skuddet. Fosfat kan reallokeres fra de eldste til de yngste delene av planten via floemet slik at fosformangelsymptomer viser seg først på de eldste bladene som visner og dør. Modning forsinkes. Planter med fosformangel kan bli rødfarget av anthocyanin analogt til nitrogenmangel. De yngste delene av planten har dypgrønn klorofyllfarge, og skiller seg således fra nitrogenmangel hvor plantene blir lysegrønne og klorotiske. Stengelen blir høy og slank, men ikke treaktig.
Litteratur
Wikipedia