{kind=link}
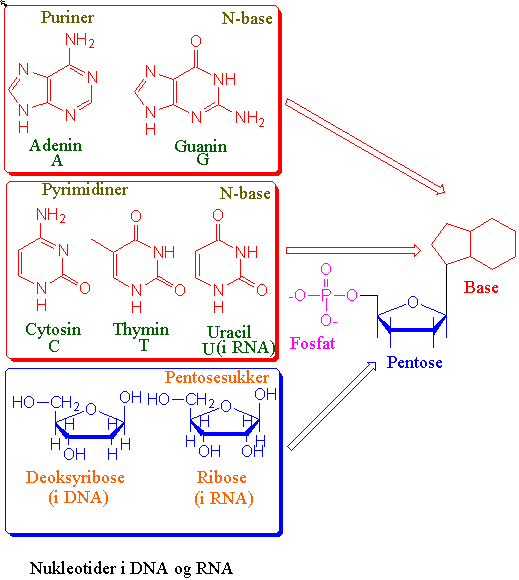
\(\displaystyle A=T\;\;\;\;\;\;\; G\equiv C\)
DNA er en stabil, men allikevel fleksibel kompakt pakket lagringsform for genetisk informasjon. Hovedstrukturen er nukleotider bundet sammen med fosfodisterbindinger. DNA er et makromolekyl med stor evne til å reparere seg selv hvis det blir skadet, og flere metoder for DNA-reparasjon. Til tross for stabiliteten har cellene stor evne til å stokke om den genetiske variasjonen via rekombinasjon og transposoner.
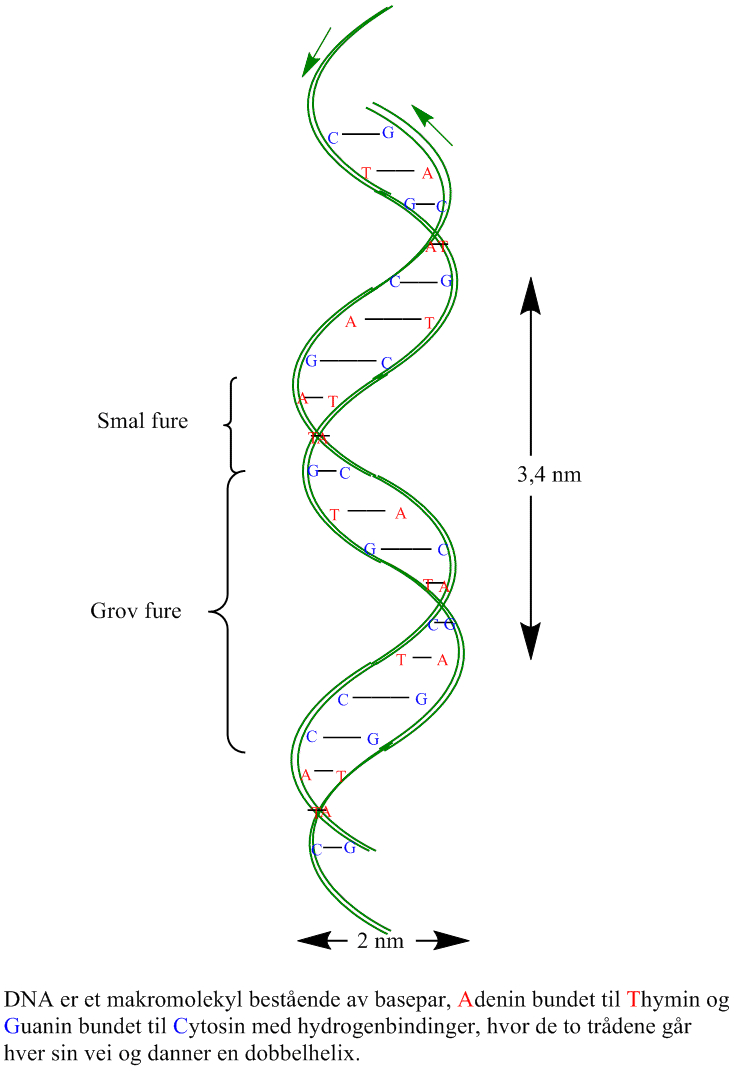
Det genetiske materialet er sammensatt av to ugreinete komplementære antiparallelle kjeder av nukleotider bundet sammen med hverandre i hydrogenbindinger som en dobbel heliks.
{kind=link}

Strukturen til DNA ble oppdaget av Watson, Crick og Rosalind Franklin. DNA-molekylet hos eukaryoter er pakket i løkker og sløyfer i flere kromosomer i cellekjernen, bundet til spesielle DNA-bindende proteiner (histoner). Komplekset mellom DNA og protein kalles kromatin, bestående av tettpakket heterokromatin som ikke blir transkribert, og eukromatin med mer åpen struktur som kan binde transkripsjonsfaktorer og bli transkribert til mRNA og deretter translatert til protein.
DNA hos prokaryoter finnes ofte i naturen som lukkete sirkulære molekyler, hvor de to enkelttrådene er bundet sammen i en sirkel. Forøvrig finnes DNA hos prokaryoter som en gonofor uten histoner (nukleoid). Kloroplaster og mitokondrier inneholder DNA med prokaryot opprinnelse i henholdsvis et kloroplastgenom og mitokondriegenom. Hos noen virus forekommer DNA som en enkelttråd, enkelttrådet DNA-virus, eller i dobbelttrådet DNA-virus.
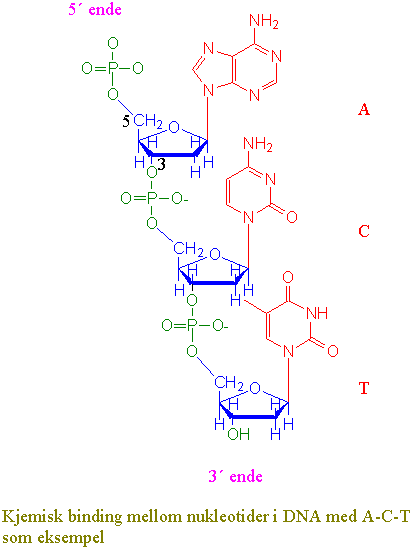
DNA er en dobbel heliks med de negativt ladete sukkerfosfatene mot utsiden og basene i par mot innsiden bundet sammen i basepar med hydrogenbindinger hvor et purin adenin bindes til et pyrimidin thymin (A-T) med 2 hydrogenbindinger og et purin guanin bindes til pyrimidin cytosin (G-C) med 3 hydrogenbindinger. Deoksyribosesukker er bundet sammen med fosfat mellom 5´-posisjon på det ene sukkeret med 3´-posisjon på det neste i en fosfodiesterbinding. Mellom trådene med nukleinsyrer tvinner det seg også en hovedfure og en bifure oppetter heliksen. De to DNA-trådene i heliksen går i motsatt retning av hverandre (antiparallelle) og de har en komplementær sekvens basesekvens. DNA i den vanlige formen (B-DNA) er en høyredreiet helix, i motsetning til venstredreiet DNA (Z-DNA). Høyredreiet vil si at heliksen dreier seg fra deg med klokken enten du ser den fra bunnen eller toppen. Det finnes også høyredreiet A-DNA i en RNA-DNA-hybrid. Z-navnet skyldes at den venstredreiete helixen får et sikk-sakk-utseende. Lokal opptvinning av helixen ved brudd av hydrogenbindingene mellom trådene tillater at RNApolymerase å lage mRNA kopier av gener og tillater DNA polymerase å replikere kopier av DNA molekylet. DNA molekylet kan tvinne seg selv og danner derved en høyere ordens heliks kalt supertvinnet DNA. Organiseringen av DNA i proteinkomplekser i kjernen gir strukturproblemer ved uttvinning av DNA trådene.
DNA kan være tvunnet i form av superkveilet DNA. Topoisomeraser er enzymer som fjerner den strukturmessige spenningen som oppstår ved uttvinningen ved å lage brudd slik at trådene får rotere fritt i forhold til hverandre for deretter å skjøte dem sammen igjen. Mye av kjerne-DNA er metylert katalysert av metylaser. Vanligvis er cytosin metylert i 5-posisjon. Man antar at metyleringen forekommer i områder som ikke deltar aktivt i uttrykk av gener. Både metyleringer og acetyleringer av histonproteiner er viktige i epigenetisk regulering av genomet.
Siden DNA er lang og tynn vil løsninger med DNA ha høy viskositet Ved oppvarming vil de to DNA-trådene atskilles i en prosess kalt DNA-denaturering (DNA-smelting). Den temperaturen hvor DNA er halvdenaturert kalles DNA-smeltetemperaturen. Senkes temperaturen og det er en relativt høy temperatur vil de to DNA-trådene gå isammen igjen (renaturering). Sirkulært DNA vris og tvinnes rundt seg selv og danner ofte superkveil. DNA er sur og negativt ladet ved nøytral pH og DNA-fragmenter vil bevege seg mot anoden (positiv elektrode) og kunne atskilles med elektroforese etter deres størrelse. DNA-fragmentene etter elektroforese vil fluorescere i UV-lys etter reaksjon med f.eks. ethidiumbromid. Tettheten til DNA varierer rundt 1.7 g cm-3 og påvirkes av innholdet av G+C. Høye konsentrasjoner av urea og formamid gir kjemisk denaturering av DNA. Basene i DNA har et absorbsjonsmaksimum ved bølgelengde 260 nanometer (nm). Ekstinksjonskoeffisienten til nukleinsyrebasene avhenger av omgivelsene, og dobbelttrådet DNA (dsDNA) her hypokrom i forhold til enkelttrådet DNA (ssDNA). Noen virus og viroider inneholder gener laget av RNA istedet for DNA. DNA er generelt mer kjemisk stabilt enn RNA.
DNA-plakking med histoner
En eukaryot celle kan inneholde fra 30.000-100.000 gener, opptil 20 x flere enn en bakterie. Dette krever pakking av DNA-tråden. DNA-heliksen tvinnes først rundt nukleosomer som ligger som perler på en snor. Nukleosomet består av histonene H2A, H2B, H3 og H4. Histonet H1 fester DNA nær "perlen". Ved hjelp av histonet H1 pakkes nukleosomene i en kromatinfiber som har en diameter på ca. 30 nm. Kromatinfiberen (30 nm) danner deretter løkker festet til et proteinstillas av ikke-histon proteiner. Dette løkkenettverket foldes og utgjør det endelige kromosomet slik det kan observeres under mitose. DNA inneholder i tillegg til de kodende sekvensene lange strekker med ikke-kodende repetert DNA og tandemrepeterte sekvenser.
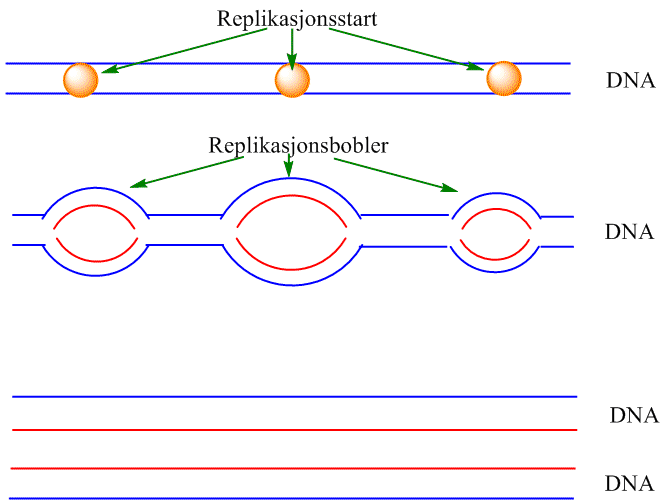
DNA-replikasjon
Replikasjon av DNA med replikasjonsbobler
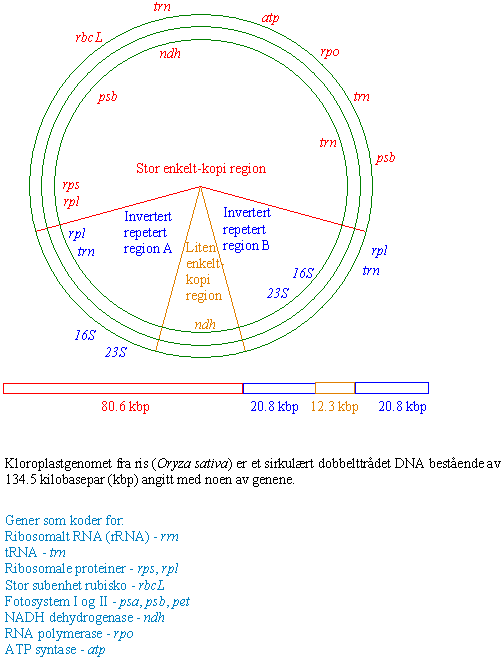
DNA i kloroplaster og mitokondrier
Kloroplast DNA, og mitokondrie-DNA behøver nødvendigvis ikke alltid være sirkulært.
Sirkulerende fritt DNA i blod
Sirkulerende (eng. circulating) fritt DNA (cfDNA) i blodplasma er blitt koblet som biomarkør till forskjellige sykdomstilstander, inkludert sirkulerende tumor DNA (ctDNA) fra kreftceller som spres i blodbaner. Sirkulerende cellefritt føtalt DNA (cffDNA) i maternalt blod med opprinnelse fra fosteret brukt i fosterdiagnostikk. Sekvenseringen av sirkulerende fritt DNA skjer via PCR, men man må som vanlig være oppmerksom på artefakter og kontaminering. Man har tilsvarende for sirkulerende RNA: cfRNA, ctRNA, cffRNA.
Litteratur
Wikipedia