Fotobiologi
Fytokrom
Lys gir ikke bare plantene energi til å drive fotosyntese og vanntransport, men kontrollerer også plantenes vekst og utvikling.
Fytokrom ble oppdaget av den amerikanske plantefysiologen L. Flint som arbeidet med effekten av lys på spiring av salatfrø, men H. Borthwick og S. Hendricks ved US Department of Agriculture, Beltsville i USA videreførte dette arbeidet som ga et gjennombrudd i fytokromforskningen.
Fytokrom er et pigmentprotein som er tilstede i alle planter i svært små konsentrasjoner. Det er lite lys som skal til for å mette de prosessene hvor fytokrom deltar. Fytokrom består av et protein bundet til en kromofor gruppe. Den kromofore lineære tetrapyrollgruppen er bundet til proteinet med en thioeterbinding. Apoproteinet lages i cellekjernen. Fykobiliproteiner som deltar i fotosyntesen hos blågrønnbakterier og rødalger har også en åpen tetrapyroll som kromofor gruppe, i motsetning til ringformet tetrapyroll i klorofyll og hem.
Fytokrom registrer informasjon om lysmiljøet omkring plantene. Fytokrom består av to interkonvertible lysabsorberende hovedformer FR og FMR. Den rødtlysabsorberende formen FR har blå farge og den mørkerødtabsorberende form FMR er grønnfarget.
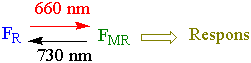
Fytokrom kan også absorbere blått lys, mens gulgrønt lys absorberes i mindre grad. Virkningen av lys som registreres av fytokrom avhenger av likevekten mellom de to formene, som igjen avhenger av forholdet mellom rødt og mørkerødt lys. De to formene av fytokrom har overlappende absorbsjonsspektere slik at det er en dynamisk likevekt mellom dem. Mengden FMR etter at fytokrom er mettet med rødt lys er ca. 85 %. Bredspektret mørkerødt lys gir ca. 97% FR og 3% FMR. Ved belysning med rødt lys skjer det en cis-trans isomeri mellom karbonatom 15 og 16. På grunn av de små mengdene med fytokrom og stort innhold av klorofyll er det vanskelig å måle fytokrom i grønt vev. Bruk av antistoffer mot fytokrom og reportergener viser at det er mye fytokrom skuddspisser, men fytokrom finnes også i rothetta.
{kind=link}
Fytokrom regulerer membranegenskapene, og rødt lys kan endre fluksen av kalsium (Ca2+) over plasmamembranen - et trinn i en molekylær signaloverføringsvei.
Plantene har flere forskjellige typer fytokromer. Fytokrom kodes av en multigenfamilie kalt PHY. De 5 forskjellige PHY-genene (PHYA til PHYE) lager fytokrom som har så forskjellig aminosyresekvens at det indikerer forskjellig opprinnelse og at de inngår i flere ulike og unike reguleringsveier. Transkripsjonen PHY-gener hemmes i lys. Fytokromprotein A er ustabilt og brytes raskt ned etter binding til ubiquitin. Ubiquitin er et polypeptid som bindes til protein og gjør det gjenkjennelig for proteosomer og nedbrytning. De fleste enzymproteiner i planter blir kontinuerlig brutt ned og bygget opp igjen avhengig av behov, og ubiquitinering gir et signal til nedbrytning.
Fysiologisk sett har plantene to typer fytokrom: type I og II fytokrom. I etiolert vev er det mest fytokrom og av type I. Det er omtrent like mye av de to formene i grønt plantevev. Mørkerødtformen (FMR) av type II er stabil i motsetning til type I PMR. Type I lages av genet PHYA og kalles nå fytokrom A. Genproduktene fra PHYB og PHYE oppfører seg som type II fytokrom.
Lys absorbert av fytokrom påvirker fotomorfogenese. Forholdet mellom rødt og mørkerødt i vanlig lys gjør at fytokromlikevekten forflyttes over mot mørkerødtformen FMR. Karakteristisk for alle fytokromresponser er at de kan reverseres flere ganger ved å gi alternerende små mengder med rødt - mørkerødt - rødt - mørkerødt osv.. Det er det lyset som gis til slutt og som flytter fytokromlikevekten den ene eller andre veien som er avgjørende. Imidlertid virker denne reverseringen bare for en kortere periode og deretter forsvinner fotoreversibiliteten.
Det mrøkerødtabsorberende fytokromformen (FMR) deltar i følgende prosesser:
- 1) Spiring hos lyskrevende frø.
- 2) Redusert strekning av internodier. Redusert strekning er også en blåttlysrespons.
- 3) Utfolding av apikal krok (hypokotylekrok) hos tofrøbladete planter når skuddet kommer opp av jorda og opp i lyset.
- 4) Redusert strekning av koleoptile og gjennombrudd av første blad hos enfrøbladete planter.
- 5) Dannelse av bladprimordier og utvikling av første blad. For å få utviklet bladplaten fullstendig trengs imidlertid høyere lysintensiteter hvor blått lys er mest effektivt.
- 6) Utrulling av blad under deetiolering. Spesielt gras har evnen til å rulle sammen bladene.
- 7) Start av kloroplastutvikling.
- 8) Vridning av kloroplasten hos grønnalgen Mougeotia.
- 9) Anthocyaninsyntese.
- 10) Turgorforandring i pulvinusceller og endring av bladvinkler. Skyldes ionefluks.
- 11) Knopphvile.
- 12) Start av redproduktiv fase.
- 13) Positiv og negativ kontroll av genekspresjon.
- 14) Regulering av cirkadiske rytmer og blomstring.
- 15) Endring i membranpotensial og fluks av ioner.
Fytokrom påvirkes av små mengder lys
Når man snakker om lysintensitet er det intensiteten av lys utsendt fra en lyskilde (sola eller kunstig belysning). Irradianse er lyset som treffer et objekt. Fluensen - antall lyspartikler som treffer en overflate, måles i mol m-2. Et mol er Avogadros tall med partikler dvs. 6.022·1023. Fluensraten eller lysfluksen er antall lyspartikler som treffer per areal- og tidsenhet og måles i mol m-2 s-1.
Det er mulig å dele fytokromresponser inn i 3 hovedtyper etter fluensrate:
1) Meget lav fluensrate (MLFR) er en effekt på fytokrom som skjer ved svært små lysmengder fra 0.1 nmol m-2 til 50 nmol m-2. Denne fytokromeffekten er ikke fotoreversibel. Det er fytokrom A som deltar og påvirkes av rødt og blått lys.
2) Lav fluensrate (LFR) er en fytokromeffekt som skjer ved lysmengder mellom 1 mmol m-2 - 1000 mmol m-2. Det er denne som er rødt-mørkerødt reversibel og det er sannsynligvis fytokrom B som deltar.
3) Høy irradianse respons (HIR) skjer i mørkespirte planter via fytokrom A. HIR responsen hos etiolerte planter har topper i mørkerødt, blått og UV A lys, og mørkerødtabsorbsjonen skyldes type I fytokrom A. Hos lysspirte planter skjer høy irradianse effekten via fytokrom B og rødt lys. HIR er ikke fotoreversibel. Typiske HIR effekter er hemming av hypokotylestrekning og fullstendig utvikling av bladplaten ved lang kontinuerlig eksponering av planten med høy irradianse.
Som et apropos til cis-trans konformasjonsendring i fytokrom skyldes synet i øyet hos vertebrater en cis-trans isomerisering i retinal. Dette gir en konformasjonsendring i rhodopsin, aktivering av G-protein, aktivering av cGMP fosfodiesterase som medfører lavere konsentrasjon av cGMP, lukking av Na+ kanaler og et elektrisk stimuli som registreres og tolkes i hjernen. G-proteiner er membranproteiner som binder GTP og GDP på en subenhet av proteinet. Hydrolyse av GTP er nødvendig for at reguleringen skal skje. Syklisk GMP (cGMP) er en sekundær budbringer som også kan delta i signaloverføringen fra fytokrom.
Fytokrommutanter og molekylærbiologi
Fytokrom regulerer genekspresjon i kjernen. Det finnes flere plantemutanter som enten har redusert mengde fytokrom eller hvor fytokromsignalveien er påvirket. For eksemple aurea (au) mutant av tomat, lang hypokotyle mutant fra agurk (lh) og lang hypokotyle mutanter (hy) av Arabidopsis. Både au og lh har redusert mengde av fytokrom type I. hy mutanter har uvirksomme gener for fytokrom og kryptokrom.
Bakterier har registreringsproteiner som består av et mottaksdomene og responsdomene. En hypotese går ut på at registreringsdomet i fytokrom er den kromofore gruppen og responsdomene er en kinase som fosforylerer proteiner.
Noen brassinosteroider deltar i lysregulerte prosesser. Steroidhormoner kan bevege seg gjennom membranenr og kan virke som transkripsjonsfaktor eller overflatereceptor.
Vegetasjonsskygge gir endret forhold mellom rødt og mørkerødt lys
Skygge forårsaket av vegetasjon gjør at forholdet mellom rødt og mørkerødt lys minsker.
Lys og frøspiring
Noen frø trenger lys for å spire. Spesielt gjelder dette planter som vokser på lysåpne steder f.eks. ugrasplanter som åkersennep, meldestokk, groblad, men også følblom, bjerk, or og furu. Høy andel med rødt i lyset fremmer frøspiring, mens høy andel mørkerødt lys hemmer spiringen. Frø av "Jomfruen i det grønne" må ha mørke for å spire, men de fleste frø spirer uavhengig av lys. Alle kulturplantene er fjernet så langt fra sin opprinnelige naturtilpassede tilværelse at disse spirer uavhengig av lys og mørke. Det er mørkerødtformen av fytokrom (FMR) som er et trinn i prosessen som får frø til å spire.
Strekning
Planter som står skyggefullt under annen vegetasjon og andre trær får flyttet fytokromlikevekten mot rødtformen av fytokrom (FR). Solplanter som står i vegetasjonsskygge blir derfor lange og strantete. Den apikale dominansen blir utpreget slik at det dannes få sideskudd. Responsen er er mest utpreget hos solplanter og observeres i liten gad hos obligate skyggeplanter.
Klatreparasitter
Lianer, vindelplanter (Convolvolus) og parasitter som neslesnyltetråd (Cuscuta europaea) har behov for å vokse inn i en vegetasjonsskygge hvor det er størst sannsynlighet for å finne en stamme eller stengel de kan klatre i (skototropisme). Fytokromsystemet har mulighet til å registrere hvor det er vegetasjonsskygge, noe som deretter påvirker vekstretningen.
Tanada effekt
Forsøk som viser at fytokrom kan gi endring i elektrisk ladning på overflaten av rotspisser av bygg. Rødt lys gjør at rotspissene fester seg til negativt ladet glass.
Blåttlysresponser
Det finnes en rekke responser i planter, alger og sopp som påvirkes av den delen av det elektromagnetiske spektrum som oppfattes som blått lys. Opprinnelig hadde man vanskeligheter med å identifisere hvilket pigmentmolekyl som registrerte blått lys og det ble en periode kalt kryptokrom - det skjulte pigment. Karakteristisk for blåttlysresponser er 3 topper i bølgelengdeområdet 400-500 nm (420 nm, 440 nm og 480 nm) pluss en topp ved 370 nm. Dette er spesifikke blåttlysresponser hvor rødt og mørkerødt lys ikke har noen effekt. Det er flere kandidater til blåttlysreceptoren: karotenoider, flaviner og pteriner bundet til protein. Lysabsorbsjonen skyldes de konjugerte dobbeltbindingene i karotenoidene og isoalloxazinringen i flavin.
Eksempler på blåttlysresponser hos planter:
- 1) Kloroplastbevegelser i cellene og fototaksis. Kloroplastene kan forflytte seg i cellene via cytoskjelettet og orientere seg til steder med optimale lysforhold.
- 2) Fototropisme - planter som bøyer seg mot lys hvor plantehormonet auxin deltar i prosessen. Fototropisme for sporangioforen, en sporebærende struktur på toppen av en lang tynn enkeltcelle, hos koblingssoppen Phycomyces bl.a. oljemugg (P. blakesleeanus).
- 3) Hemming av hypokotylestrekning. Lys reduserer strekning.
- 4) Økt syntese av klorofyll og karotenoider. Blåttlyseffekt på genet som koder for enzymet glutamat-1-semialdehyd aminotransferase i klorofyllbiosyntesen hos grønnalgen Chlamydomonas reinhardtii.
- 5) Åpning av spalteåpninger (stomata). Stomata åpnes ved en blåttlysaktivert H+-ATPase i lukkecellenes plasmamembran. Man må skille mellom åpningen som skyldes fotosyntese og den rene blåttlysresponsen. Eksperimentelt kan man bruke rødt lys til å mette fotosyntesen og gis deretter blått lys får man større åpning.
- 6) Fotoreaktivering og reparasjon etter UV-skader. Fotoreaktivering er en enzymatisk reparasjonsprosess hvor en fotolyase deltar. Det blåttlysabsorberende enzymet binder seg spesifikt til syklobutyl-pyrimidin dimere i DNA, forårsaket av UV-lys, og reparerer skaden. Derfor repareres UV-skadene raskere i lys enn i mørke.
- 7) Økt respirasjon.