- Hva er fett ?
- Fett som opplagsnæring i frø
- Spiring av fettfrø - fra fett til sukker
- Fettsyresyntese
- Hvordan lages glycerol-3-fosfat og veien videre til fett
- Galaktolipider MGDG og DGDG, fosfolipider og sulfolipider
- Flytende planteoljer
- Oksidasjon av fettsyrer
Hva er fett ?
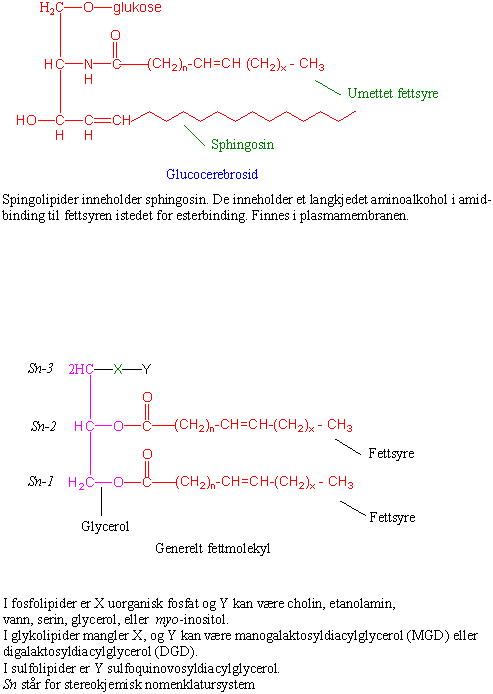
Acyl henspeiler på fettsyrene. Fett kalles olje når det er flytende ved romtemperatur og dette skyldes høyt innhold av umettede fettsyrer i fettet. Fett som er i fast form ved romtemperatur inneholder mer mettede fettsyrer. Fett er bygget opp av 3- karbonforbindelsen glycerol med 3 hydroksylgrupper (-OH) hvor 2 eller 3 fettsyrer er festet med esterbinding mellom syregruppene i fettsyrene og hydroksylgruppene i glycerol. Det finnes mange forskjellige fettsyrer i planter. I fosfolipidene er det festet fettsyrer til to av hydroksylgruppene i glycerol ("fettsyrehalene"), mens til det tredje hydroksylgruppen er det festet et "hode" med enten fosfatidylcholin (PO32--CH2-CH2- N+(CH3)3), fosfatidinsyre (HPO32-), fosfatidyletanolamin (PO32--CH2-CH2-NH2), hydrogen (-H; diacylglycerol) eller galaktose (galaktolipid). Fosfolipider er polare og en viktig bestanddel av biologiske membraner. Egenskapene til membraner påvirkes av hvilke fettsyrer som inngår og i hvilken grad de er mettet eller umettet. Jo mer umettet fett desto mer flytende egenskaper i membranen. Fosfolipidene er diglycerider. Den hydrofobe (vannskyende) delen med fettsyrene vender inn i membranen og den hydrofile (vannelskende) polare delen av fettmolekylet vender ut mot cytoplasma. Et molekyl som har både hydrofile og hydrofobe egenskaper kalles amfipatisk. Det er forskjellige typer fosfolipider avhengig av hvilke typer fettsyrer og fosforgrupper som inngår i molekylet. Lipider som inneholder inneholder sukker kalles glykolipider. Det finnes også sulfolipider og sphingolipider. Kloroplastmembranen inneholder omtrent 70 % glykolipider (monogalaktosyldiacylglycerol (MGDG) og digalaktosyldiacylglycerol (DGDG), og ca. 7 % fosfolipider og 7 % sulfolipider.
{kind=link}
{kind=link}
De vanligste mettede fettsyrene i planter er laurinsyre (12:0), myristinsyre (14:0), palmitinsyre (16:0) og stearinsyre (18:0). Antall karbonatomer og antall dobbeltbindinger mellom karbonatomer er angitt i parentes. De vanligste umettede fettsyrene i planter er oljesyre (18:1), linolsyre (18:2) og linolensyre (18:3). Andre fettsyrer som finnes i planter er erukasyre hos korsblomster, ricinoleinsyre hos Ricinus og arachidonsyre (20:4). Noen plantefamilier inneholder spesielt mye C8- C14-fettsyrer i fettet. Arter i almefamilien (Ulmaceae) kan inneholde mye fettsyrer av typen 8:0 og 10:0; laurbærfamilien (Lauraceae) med 10:0 og 12:0; og muskatnøttfamilien (Myristicaceae) med 14:0. Palmer i palmefamilien (Arecaceae) har stor variasjon i fettsyreinnhold.

Fett som opplagsnæring i frø
Fett inneholder mye karbon og hydrogen i forhold til oksygen, det vil si mye potensiell energi som kan frigis ved oksidasjon. Triglycerider er den vanligste lagringsformen av fett. Noen frø inneholder relativt mye fett (solsikke, gran, sojabønner, raps, kokosnøtt, bomull, og lin). Fettet er lagret i endosperm eller frøblad. Avokado og oliven er eksempler på frukt som inneholder mye fett. Fett (lipider) finnes som opplagsnæring i organeller kalt oleosomer i frø. Oleosomene er omgitt av en membran med bare den ene halvdelen av dobbeltlaget med fosfolipider. Den hydrofobe delen vender inn mot fettet og den hydrofile delen ut mot cytoplasma er stabilisert med spesielle proteiner kalt oleosiner.
Spiring av fettfrø - fra fett til sukker
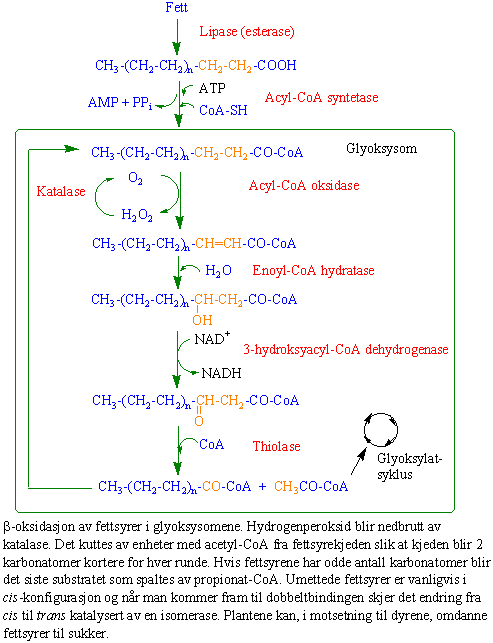
Fett lagret i oleosomer i frø blir omsatt under frøspiringen. Først brytes hydrolyttisk bindingen mellom fettsyrene og glycerol katalysert av enzymet lipase. De frie fettsyrene fraktes inn i glyoksysomer hvor de oksideres ved -oksidasjonb. Her spaltes det av to og to karbonatomer i form av acetyl-CoA. Acetyl-Co inngår i glyoksylatsyklus. Først reagerer acetyl-CoA med oksaledikksyre og danner citrat katalysert av enzymet citrat syntase. Citrat vil ved isomerisering danne isocitrat. Deretter spaltes isocitrat katalysert av isocitrat lyase til succinat og glyoksylat. Succinat overføres til mitokondriene og omdannes til malat (eplesyre) ved å bruke en del av trikarboksylsyresyklus. Malat fraktes deretter ut i cytosol og enzymet malat dehydrogenase omdanner malat til oksaledikksyre. Oksaledikksyre danner fosfoenolpyruvat katalysert i en ATP- avhengig dekarboksyleringsreaksjon av enzymet fosfoenolpyruvat karboksykinase. Fosfoenolpyruvat kan omsettes ved reversering av glykolysen (glukoneogenesen) til sukker. Hva med glykosylat ? Jo, den reagerer med nok et molekyl acetyl-CoA fra fettsyrene og omdannes til malat katalysert av malat syntetase. Deretter lages oksaledikksyre fra malat katalysert av malat dehydrogenase og oksaleddiksyre er klar for å reagere på nytt med et molekyl acetyl-CoA.

Fettsyresyntese
Syntesen av fett skjer i samarbeid mellom kloroplaster og endoplasmatisk retikulum. De siste trinnene i syntesen av triglycerider skjer i endoplasmatisk retikulum og fettet akkumuleres mellom de to dobbeltlagene i enhetsmembranen.
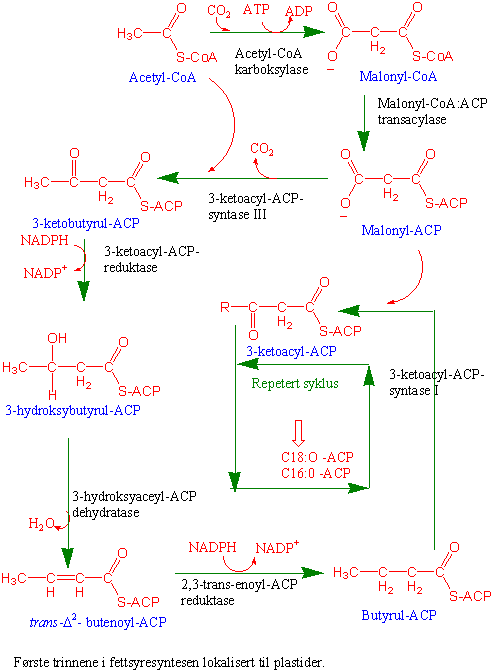
Kloroplastene er hovedsted for fettsyresyntese i blad med endeproduktene palmitinsyre og oljesyre. Disse fraktes deretter ut i cytosol hvor de omdannes videre til linolsyre og linolensyre. Utgangsmaterialet for biosyntese av fettsyrene er tokarbonthioesteren acetyl CoA, laget fra importert acetat. Lavmolekylært ACP ("acyl carrier protein") hjelper til å koble malonyl-ACP til å forlenge acylkjeden. Acyl er festet kovalent til ACP via en thioesterbinding
Fettsyresyntesen skjer i et stort enzymkompleks kalt fettsyre syntase. Første trinnet er at acetyl- CoA omdannes til malonyl-CoA i en ATP-avhengig karboksylering katalysert av acetyl-CoA karboksylase som er et av enzymene i komplekset fettsyre syntase. Malonylgruppen i malonyl-CoA overføres til et lavmolekylært surt acylbærer protein kalt ACP ("acyl carrier protein") og det dannes malonyl-ACP katalysert av malonyl-CoA:ACP transacylase. Acylkjeden er bundet med en thioesterbinding. Fra nå av skal det hektes på tokarbonenheter fra acetyl-CoA. Først overføres acetat fra acetyl-CoA til et kondenserende enzym kalt 3-ketoacyl-ACP syntase. Forlengelsen av fettsyrene katalyseres av de fire enzymene 3-ketoacyl-ACP syntase, 3-ketoacyl- ACP reduktase, 3-hydroksyacyl-ACP dehydrase og enoyl-ACP reduktase. Ketogruppen ved karbonatom 3 fjernes som karbondioksid. Det skjer en reduksjon av 3-ketogruppen katalysert av 3- ketoacyl-ACP reduktase og det dannes 3-hydroksybutyrul-ACP. Deretter skjer det en dehydrering katalysert av hydroksyacyl-ACP dehydratase og det dannes trans-D2-butenoyl-ACP. Ved en reduksjon av dobbeltbindingen katalysert av enoyl-ACP reduktase blir produktet butyryl-ACP med 4 karbonatomer.
{kind=link}
![]()
Deretter går butyrul-ACP inn i ny syklus hvor det på nytt hektes på to karbonatomer fra malonyl-ACP katalysert av 3-ketoacyl-ACP syntase inntil kjeden består av 16 (palmitoyl-ACP) eller 18 (stearoyl-ACP) karbonatomer.
![]()
De fleste umettede fettsyrene inneholder en dobbeltbinding ved karbonatom 9. Stearoyl-ACP desaturase er et enzym som finnes i stroma i plastidene, trenger oksygen og bruker redusert ferredoksin eller NADPH som elektronkilde. Dette enzymet lager den første cis-dobbeltbindingen i fettsyrene, mellom karbon 9 og 10, og produktet blir oleoyl-ACP. Petroselinsyre, som er en fettsyre det er mye av i frø fra skjermplantefamilien, er imidlertid en C18:1cisD6 fettsyre. Selv om fettsyresyntesen skjer i plastider er de fleste genene som koder for enzymene i biosynteseveien lokalisert til cellekjernen. Flere av enzymene som deltar i fettsyresyntesen finnes som isoenzymer og genene for disse finnes i multigenfamilier. Enzymet acyl-ACP thioesterase som fjerner ACP fra fettsyrene er med å bestemme hvor lange fettsyrene skal bli.
{kind=link}
Hvordan lages glycerol-3-fosfat og veien videre til fett
Endring av acylkjeden via hydroksylering, avmetning og forlengelse skjer utenfor plastidene. Syntese av fett (triglycerider) skjer via Kennedy-biosynteseveien på endoplasmatisk retikulum. Fosfat fjernes fra fosfatidinsyre og gir diacylglycerol som danner et greiningspunkt mellom dannelse av fosfatidylcholin i membranene og triglycerider for lagring av fett. Glycerol-3-fosfat, som er den aktive formen av glycerol i syntese av lipider, lages fra dihydroksyacetonfosfat katalysert av enzymet dihydroksyacetonfosfat reduktase. Det er også en mulighet for at glyceraldehyd-3-fosfat kan defosforyleres til glycerol katalysert av defosforylase reduktase. Deretter kan glycerol-3-fosfat lages med hjelp av enzymet glycerol kinase. Sukker til galaktolipidene kommer via UDP-galaktose.
Neste trinn for å lage triglycerider er to acyleringsreaksjoner hvor fettsyrene bundet til ACP reagerer med glycerol-3-fosfat. Acylering av glycerol-3-fosfat skjer inne i plastider fra fettsyrene som CoA-estere (palmitoyl-ACP og oleoyl-ACP). I kloroplaster lages lipidene, kalt prokaryote lipider, ved overføring av acylgruppen fra ACP til glycerol-3-fosfat i en esterifiseringsreaksjon i Sn1-posisjon katalysert av glycerol-3-fosfat-acyltransferase. I denne reaksjonen overføres oljesyre til Sn1-posisjon i glycerol. Det dannes lyso-fosfatidinsyre (Sn1-1-oleoyl-glycerol-3-fosfat). I neste trinn katalysert av lysofosfatidinsyre acyltransferase (1-acylgylcerol-3-fosfat acyltransferase) lages fosfatidinsyre (1,2-diacyl-glycerol-3-fosfat). Siden Sn-1-posisjonen allerede er opptatt plasseres som oftest palmitinsyre (C16:0) i Sn-2-posisjon. Esterifisering av Sn3 katalyseres av en diacylglycerol acyltransferase.
Fosfatidinsyre kan enten brukes til å lage fosfatidylglycerol eller diacylglycerol. Første trinn på veien til fosfatidylglycerol er en reaksjon mellom fosfatidinsyre og CTP katalysert av fosfatidat cytidyltransferase og det dannes CDP-diacylglycerol. Deretter hektes det på glycerol-3-fosfat i Sn- 3-posisjon katalysert av CDP-diacylglycerol-glycerol-3-fosfat 3-fosfatidyltransferase. En fosfatidylglycerofosfatase fjerner fosfat og den desaturase kan lage fettsyrene mer umettet.
Første trinn på veien til diacylglycerol er fjerning av fosfat fra fosfatidinsyre katalysert av fosfatidat fosfohydrolase. Alternativt kan det henges på sukker fra UDP-galaktose i Sn-3-posisjon katalysert av MGDG syntase for å lage glykolipider eller UDP-sulfoquinovose katalysert av sulfolipid syntase for å lage sulfolipider. Lipider som lages i cytoplasma kalles eukaryote lipider fordi det har en fettsyresammensetning som man finner hos dyr.
Galaktolipider MGDG og DGDG, fosfolipider og sulfolipider
Galaktolipider lages av enzymer i kloroplastmembranen fra glycerol-3-fosfat fra cytoplasma som sammen med Acyl-ACP (ACP- "acyl carrier protein") gir lysofosfatidinsyre
![]()
som igjen binder acyl-ACP og gir fosfatidinsyre.
![]()
Fosfatidinsyre omdannes til diacylglycerol
katalysert av UDP-galaktose diacylglycerol galaktosyltransferase.
![]()
Neste enzym danner slutten UDP-galaktose-MGDG galaktosyltransferase
eller galaktolipid galaktosyltransferase
![]()
Flytende planteoljer
Planteoljene er flytende ved romtemperatur på samme måte som olje fra marine organismer. Planteoljer fås fra frø som har mye fett som opplagsnæring slik som solsikke, soyabønner, mais, kokosnøtter, raps og lin. Fettet kan presses ut av frøene. De viktigste umettede fettsyrene i planteoljer er oljesyre, linolsyre og linolensyre og fett med disse har vist seg å være gunstige ernæringmessig i forhold til mettede fettsyrer fra dyreriket. Jo kaldere en plante vokser desto mer umettede fettsyrer inneholder membranene. Dette er en måte planten kan holde membranene mest mulig lettflytende ved lav temperatur. Fettsyrene fra planteriket er ikke bare uskyldige. Opptil 50 % av fettet i noen eldre varieteter av raps inneholder erukasyre. Dette er en umettet fettsyre som gir"fetthjerte" hos forsøksdyr. Sterculinsyre er en giftig syklisk fettsyre.
Helseskader som følge av petroleumsbaserte oljer har ført til økt interesse for bruk av planteoljer til industriformål f.eks. til skjæreoljer. I behandling av treverk har linolje vært mye brukt. Linolje tørker i luften når fettsyrene reagerer med oksygen i lufta. Kokt linolje, som er linolje kokt med blyoksid, tørker fortere enn den rå linoljen. Ekte linoleum er et gulvbelegg som er laget av linolje og presset og festet på et nettverksunderlag av jute. Planteoljene er også brukt i hudkrem.
Oksidasjon av fettsyrer
Fettsyrer kan oksideres ved beta-oksidasjon i glyoksysomene eller ved alfa-oksidasjon.
{kind=link}
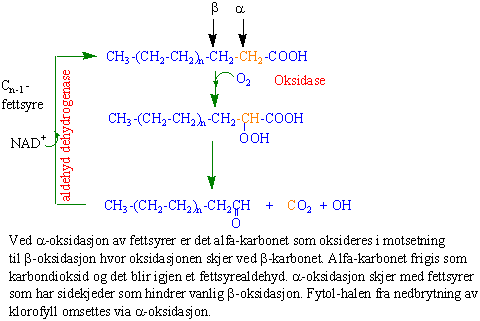{kind=link}
Linolensyre brukes også som utgangsmateriale for å lage plantehormonet jasmonat. Hydroperoksider av de umettede fettsyrene linolsyre og linolensyre lages i en reaksjon katalysert av lipoksygenase. Lipoksygenaser er ikke-hem jernproteiner som oksiderer fettsyrer som inneholder et sete med cis-cis-1,4-pentadien. Lipoksygenasene forekommer som isoenzymer og har flere funksjoner i plantene.