Genmodifiserte planter
Hvorfor brukes antiobiotikaresistensgener ?
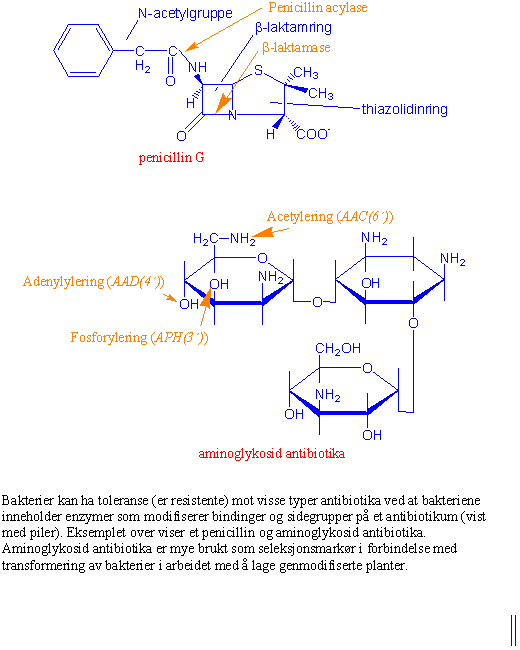
Det fremmede genet (transgenet) settes bare inn i noen få celler. For å kunne identifisere og finne igjen hvilke celler eller planter som inneholder det nyinnsatte genet overfører man samtidig et markørgen f.eks. for antibiotikaresistens. For å kunne kontrollere at genet er satt inn på riktig sted og at det blir uttrykt overfører man markørgener sammen med genet som skal overføres. Et vanlig markørgen er neomycin fosfotransferase (nptII) som koder for enzymet aminoglykosid-3´-fosfotransferase II (APH(3´)II). Dette enzymet gir en ATP- avhengig fosforylering av 3´-hydroksylgruppen på aminoheksoseringen. Aminoglykosidantibiotika som kanamycin, neomycin og streptomycin binder seg til ribosomene og blokkerer translasjon hos bakterier, men hemmer også proteinsyntesen i kloroplaster og mitokondrier. Celler eller planter som mangler markørgenet vil ikke vokse og celledlingen stopper. Plantedeler som inneholder markørgenet blir resistente eller tolerante mot det utvalgte antibiotika.
Om antibiotika
Antibiotika produseres i lave konsentrasjoner i mikroorganismer og brukes av disse for å hindre vekst av bakterier. Viktige produsenter av antibiotika er aktinobakterier (tidligere kalt aktinomyceter). Dette er filamentformete bakterier som lever i jord og vann hvor de bryter ned organisk stoff. De trådlignende aktinobakteriene gir lukt av jord. Jordmikrobiologen Selman Abraham Waksman (1888-1973) fikk nobelprisen i fysiologi og medisin i 1952 for oppdagelsen av streptomycin, produsert av aktinomyceten Streptomyces griseus Streptomycin kunne brukes i behandlingen av tuberkulose.
Waksman oppdaget også et antibiotika kalt neomycin. Både streptomycin, neomycin og kanamycin er aminoglykosider som hemmer proteinsyntese i bakterier, og lages av arter av Streptomyces. Flere typer antibiotika binder seg til eller hemmer proteiner i bakterier. Penicilliner og cephalosporiner har en b-laktamring som påvirker proteiner bakteriene bruker til å lage cellevegger. Polymyxin og colistin er antibiotika som binder seg til fosfolipider i membranen og gir økt lekkasje av stoffer ut gjennom bakteriemembranene. Rifampin reduserer syntesen av RNA ved å binde seg til enzymet som kopierer RNA. Tetracykliner er bredspektret antibiotika som påvirker proteiner og hemmer bakterier som er i aktiv deling. Kloramfenikol som binder seg til 70S ribosomer i bakterier og hemmer proteinsyntesen ble opprinnelig funnet i Streptomyces venezuelae.
Andre seleksjonsmarkører
Den motstanden bruk av antibiotikaresistensgener skapte i opinionen har gjort at molekylærbiologiindustrien forsøker å bruke andre seleksjonsmarkører, eller å fjerne seleksjonsmarkøren etter bruk.
Bioluminiscens
En mye brukt seleksjonsmarkør er enzymet luciferase som kodes av genet luc. Luciferase vil sammen med det organiske molekylet luciferin gi utsendelse av lys i nærvær av oksygen.
Luciferin fra ildfluer er forskjellig fra det man finner i bakterier med bioluminiscens. Hos ildfluer reagerer ATP, magnesium (Mg2+), luciferase og luciferin i et luciferase-luciferyl- adenylat kompleks som sammen med oksygen gir utsendelse av lys. Ildfluehannen sender under flyving ut pulser med lys som registreres av hunner på bakken. Hunnen svarer med et lysglimt og derved tiltrekker hannene. Hos bakterier skjer lysutsendelsen ved enzymatisk oksidasjon av redusert flavinmononukleotid (FMNH2). Luciferase, FMNH2 og oksygen reagerer med langkjedete alifatiske aldehyder som gir lysutsendelse. Det finnes mange andre organismer som har bioluminiscens. Dinoflagellater, fisk (Apogon, Parapriaeanthus) med lys fra fotocyster, St.-Hans orm, muslingkreps (Cypridina hilgendorfii) og maneter.
Grønt fluorescerende protein (GFP) er en annen markør. GFP er proteinet aequorin fra en manet (Aequorea) som gir bioluminiscens i nærvær av kalsium- eller strontiumioner. I dette tilfellet trengs ikke andre oksidasjonsmidler.
Andre markører
Fosfomannose isomerase kodet av genet manA/pmi har også blitt brukt. Planteceller med markørgenet pmi kan omdanne mannose-6-fosfat (som planten lager fra mannose katalysert av heksokinase) til fruktose-6-fosfat, og unngår derved de toksiske effektene. Et annet markørsystem basert på samme prinsipp er xylose isomerase (xylA) som gjør at plantevevet kan vokse på xylose. Genet ipt koder for isopentenyl transferase som lager isopentenyl- AMP som omdannes til cytokinin. Dette markørgenet gir kallus på hormonfritt medium. Plantecellekulturer som vokser i et medium tilsatt mye av aminosyrene lysin og threonin vil ikke vokse pga. mangel på methionin. Feedbackufølsom aspartat kinase kan i et slikt tilfelle brukes som markørgen.
,