Kjøleskader og frostskader
"Kvar vart det taa alle dei blomar, som stod og nikka i ljos og i ange ?" Per Sivle "Haust"
- Herdingsprosess og kuldeaklimatisering
- Underkjølt vann i frostherdete urter, busker og trær
- Membraner og lav temperatur
- Isolerende snøleier
- Skoggrense og temperatur
- Naturkalender
- Høstfarger
- Temperatur og temperaturmåling
Det er bare ca. 7 % av jorda i et belte rundt ekvator hvor temperaturen ikke synker under 15oC i løpet av året: Amazonas, Sentral-Afrika og Sydøst-Asia - et område som følger utbredelsen av palmer. I Sibir, Alaska og Nord-Canada kan temperaturen bli lavere enn - 50oC, og her kan trærne kan stå 3-4 måneder i frossen tilstand. Meteorologiske data som tas to meter over bakken gir ikke alltid beskrivelse av hvor planter utsettes for frost. Det er to hovedtyper skader av lav temperatur:
1) Kjøleskader. Planter kan få skader av lav temperatur mye før temperaturen nærmer seg frysepunktet for vann. Særlig tropiske planter som er tilpasset å vokse på steder med høy temperatur kan få kjøleskader. Disse opptrer når temperaturen er mellom 2 og 13oC, det vil si før vann fryser. Kjøleskadene gjør at membranegenskapene ødelegges. Banan som har ligget i kjøleskapet vil vise kjøleskader. Følsomheten for kjøleskader ser ut til å være korrelert med hvilken grad det er umettede fettsyrer i fosfatidylglycerol i kloroplastmembranene. Planter med høyt innhold av cis-umettede fettsyrer er resistente mot kjøleskader. Kjøleskader har vært koblet til faseoverganger i membraner fra væskekrystallinsk tilstand til en gelform.
2) Frostskader (fryseskader) oppstår ved temperatur lavere enn 0oC, og skyldes iskrystaller i cytoplasma. Som følge av konveksjon vil plantenes temperatur nå lufttemperatur i løpet av kort tid hvis de ikke utsettes for direkte solstråling. Massive plantedeler med mye vann synker ikke så raskt i temperatur som de med lite vann. Varmeoverføring skjer raskere ved bladspisser og kanten av blad, og gir et karakteristisk mønster for frostskadene. Langs kantene og spissen av blad blir det lettest rimdannelse en frostnatt. Plantene kan velge to overlevelsestrategier: Tåle frosten eller unngå den. Mange planter dør av frostskader hvis temperaturen blir lavere enn - 1 - -5 oC, mens de første vårplantene tåler godt slike temperaturer. Følsomheten for frost varierer med årstiden, og plantene er minst følsomme for frost vinter, vår og høst. Bartrær som er herdet tåler 30-40 kuldegrader uten å ta skade. Hos mange planter dør de overjordiske delene den første frostnatten om høsten. Plantene kan unngå problemene med lav temperatur ved å beskytte seg med en spesiell vokseform eller ha en lissyklus og utvikling som unngår eksponeringen for den lave temperaturen. Mange to og flerårige planter unngår vinterkulden ved å danne overvintringsorganer som rotknoller, jordstengler, løk eller frø. Både potetplanten og georginer fryser ved 0oC og blir misfargede, væskefylte og slappe, for deretter å bli brune. Dette skyldes ødeleggelse av membraner. Lagres poteter under 0oC omdannes stivelse til sukker. Vannet i plantene fryser ved temperaturer lavere enn 0oC hvis de er kuldeherdet. Frostskader kan gi korkstriper på frukt. Vann utvider seg når det fryser og lager iskrystaller. Utvidelsen kan gi frostsprekker på stamme og greiner hos trær. Spesielt utsatt er frukttrær og varmekjære løvtrær som spisslønn (Acer platanoides), eik (Quercus), ask (Fraxinus excelsior) og alm (Ulmus glabra). Bredden på frostsprekkene er sterkt korrelert med temperaturen i de ytre delene av stammen. Frost kan gi forsinket utvikling av knopper, særlig blomsterknoppene er følsomme for lav temperatur. "Solavskalling" skyldes frysing og finnes vesentlig på den syd-sydvestre delen av trestammer opptil 2 m over bakken. Den nedre grensen avhenger av snødybden. I alvorlige tilfeller skilles kambiet fra veden. Solavskallingen skyldes alternerende frysing og tining. Soloppvarming av trestammen kan gi temperaturøkning på opptil 20 oC i forhold til skyggesiden. Når sola plutselig forsvinner faller temperaturen raskt på de solbestrålte områdene. Bladfelling hos løvtrær er en morfologisk tilpasning til lang vinter, noe som reduserer vanntapet . Klorofyll i innerbarken hos trær og busker kan kompensere for en lang bladløs sesong. Blåbær (Vaccinium myrtillus) har en vintergrønn stengel som kan fotosyntetisere. Fotosyntese i den grønne innerbarken kan lage sukker som senker vannpotensialet i floemcellene. Tykk bark på trestammer virker isolerende. Det er gunstig med stort volum i forhold til overflaten. Kaktus og stammesukkulenter benytter seg av dette. Det indre av en tue- eller mattedannende plante kan være flere grader varmere enn omgivelsene. Tuer og matter kan motta mye jordvarme fra omgivelsene og er en vanlig vekstform hos fjellplanter. Krystalliseringsvarmen for vann gir varmeproduksjon ved frysing. Det kan ta flere timer før et vannmettet strøfall fryser. Moser tåler godt frysing. Vannet fryser utenpå mosene og mosene har liten intern vanntransport. Vannet beveger seg via kapillarkrefter oven- og nedenfra. Moser danner ofte tette matter og vannet går rett inn i bladet. Moser tørker lett ut, men om nettene reduseres vanntapet i forhold til et enkeltindivid.

Frostskadet bringebær om våren

Frostskadet geitrams. Bladene blir hvite av fotooksidert klorofyll.

Frostskadet bregne om våren

Frostskadet fugletelg.

Frostskadete blader hos osp om våren.

Frostskadet kålplante på Østlandet i juni.
Bartrærne er godt tilpasset lav temperatur, selv om nålene sitter på om vinteren. Nålefellende bartre som lerk (Larix) kombinerer begge vekstformer. Hovedproblemet om vinteren er lav temperatur og uttørking. Trærne kan akklimatiseres til å tåle temperaturerer under 0oC hvor vannet i levende celler underkjøles. Planter som er herdet til å tåle frost har evnen til å lage is i cellevegger og intercellularrom (apoplast), mens symplasten holdes underkjølt og fri for is. Ekstracellulær frysing skjer i intercellularrom, på overflaten av cellene eller mellom protoplast og cellevegg. At is først dannes utenfor cellene er viktig for overlevelsen. Iskrystaller som utvider seg inne i cellene ødelegger membranene. Iskrystallene kan imidlertid vokse mellom cellene uten å gi skade. Plasmamembranen kan skyves innover i cellen av iskrystallene uten at disse trenger igjennom. Vanndamptrykket for vann i cellene er høyere enn for is ved samme temperatur. Vann vil derfor fortsette å diffundere gjennom plasmamembranen til den ekstracellulære isen. Vanndamptrykket er et uttrykk for mengden vanndamp som er i likevekt med flytende vann eller is. Vann beveger seg fra et underkjølt sted på innsiden av cellen til utsiden. Dette gir en dehydrering av cellen. Forskjellen i kjemisk potensial mellom vann i underkjølt tilstand og det kjemiske potensialet til den ekstracellulære isen betyr at cellen tørker ut. Is har lavere vanndamptrykk og kjemisk potensial enn væske ved samme temperatur, og vann vil derfor vandre ut av cellene og ut til den intercellulære isen. Frostherdige planter har evnen til å rehydrere cellene etter dehydreringen. Når vann beveger seg ut av cellene vil celleinnholdet skrumpe inn. Dette betyr at frysepunktet blir lavere for det innholdet som er igjen i cellene. Stiger temperaturen går vannet tilbake til plantecellen hos herdige planter. Man kan se hvordan dette fungerer hos Rhododendron hvor bladene henger vertikalt sammenrullet ved ca. -10 oC, men retter seg opp ved ca. -3oC. Dette skyldes vann som vandrer ut og inn av plantecellene avhengig av temperaturen.Cellenes sukkerinnhold kan også øke ved lav temperatur,slik man har hos poteter som blir lagret ved kuldegrader. Sukkerinnholdet gjør at vannet i cellene ikke fryser så lett. Intracellulær frysing skjer når temperaturen kommer under 0oC, og cellene ødelegges av iskrystallene. Frysingen skjer kontinuerlig hvis vevet er homogent f.eks. parenkym i blader. Frysingen kan også skje sekvensielt hvis f.eks. ledningsvevet fryser først og deretter parenkymet. Diskontinuerlig frysing skjer i større trær og greiner. Barkparenkym kan fryse ved ca. -10oC, mens vedparenkym kan underkjøles til en lavere temperatur før de fryser gruppevis. Is dannes først i de brede vedrørene hvor det er vanskelig å få underkjølt vannet. Vann som fryser i xylemceller vil gi kavitering. Luftbobler som har blitt frosset ut vil stå under trykk hvis linseporene er lukket. Ved tining løses luftboblene i xylemvæsken. Bartrærne har bare trakeider, og med smalere diameter er trakeidene mindre utsatt for kavitering, sammenlignet med angiospermene som har vide vedrørselementer i tillegg til trakeidene. Når vann fryser i vedrørene vil ufrosset vann strømme vekk fra froststedet og det vil ikke bygge seg opp noe trykk som kan løse luftbobler. Saftstigning i løvtrær om våren ved rottrykk eller stammetrykk kan delta i reparasjon av vedvev som er blokkert ved kavitering. Når en stamme eller grein nedkjøles synker temperaturen i lufta og stammen ned til ca. -8 - - 10oC. Hvis lufttemperaturen fortsetter å synke kan temperaturen i stammen stige til 0oC. Dette er første eksoterm hvor vannet utenfor cellene fryser til is og frigir varme. Første eksoterm kan ha to topper. Den ene skyldes vann som fryser i veden (xylem), og den andre vann som fryser i mikroporer og cellevegger. Frost tidlig i vekstsesongen er mye mer skadelig enn om vinteren. Andre eksoterm som kommer ved ca. -35 til -40oC skyldes intracellulær frysing av vann som frigis fra makromolekyler.
For å få underkjølt planteceller må følgende betingelser være oppfylt:
- 1) Liten cellestørrelse.
- 2) Få eller ingen intercellularrom for iskjernedannelse.
- 3) Relativt lavt vanninnhold .
- 4) Fravær av indre iskjernedannere.
- 5) Nærvær av antiiskjernedannere.
Plantecellene kan overleve nedkjøling i flytende nitrogen hvis vanninnholdet er lavt og hvis kjølingen og oppvarmingen går meget raskt. Tørre plantedeler har noe vann bundet i matriks, men dette blir ikke omdannet til is når plantene kjøles ned til meget lave temperaturer. Bregneblad som f.eks. skades ved -10 - -20oC når de er mettet med vann, tåler -50oC når de har 5% vanninnhold. Kritisk vanninnhold for å overleve nedkjøling i flytende nitrogen (- 196oC) er fra 15-20% av friskvekt. Plantene kan overleve nedkjøling i flytende nitrogen hvor vann går over i fast form uten at vannmolekylene reorienteres til iskrystaller. Prosessen kalles vitrifikasjon,og det skjer i en amorf stivning av celleinnholdet (glassformasjon). Tørket frø kan beholde spireevnen i mange år hvis det lagres ved ca. -20oC.
Når vann i bakken og i ledningsvevet er frosset og plantene fortsetter å transpirere blir resultatet vintertørke. Eviggrønne planter kan utvide vekstsesongen noe i forhold til løvfellende trær og busker, men fotosyntesen er lav ved lav temperatur. Kuldegrader og sterkt sollys utsetter fotosynteseapparatet hos barnåler for lystress siden de biokjemiske CO2- reduserende enzymsystemene er satt ut av drift ved den lave temperaturen.

Iskrystaller på overflaten av vintergrønne tyttebærblad.
"Det blomstrer til sneen faller og dekker busk og strå". Arnulf Øverland "Denne vår jord"
Herdingsprosess og kuldeaklimatisering
Før vinteren må løvtrærne kvitte seg med bladene tilknyttet treets vannledningssystem. Da trærne ikke kan få tak i vann fra den frosne jorda ville vanntap fra bladene ha ført til tørkeskader. Bare trær med tørketilpassede blad kan ha bladene hengende på om vinteren f.eks. barnåler hos gran (Picea abies), furu (Pinus sylvestris) og einer (Juniperus communis), og tyttebær (Vaccinium vitis-idaea) og Rhododendron. For å kunne overleve vinteren må plantene være istand til å tåle at vannet fryser til is. At vannet virkelig er frosset til is i grener og stammer vet alle som har hugget trær en kald vinter. Når nettene blir lenger på ettersommeren går de flerårige trær og busker inn i vekstavslutning og blir kuldeherdet. Kuldeherdete planter overlever lav temperatur og unngår frostskader. Kuldeherdigheten består i endring av membranegenskaper og proteininnhold slik at cellene tåler uttørking, og slik at cellene kan ta opp vann når isen tiner i intercellularrommene. Deretter gjenoppretter cellene livsaktiviteten. Faktorer som øker veksten f.eks. sterk nitrogen gjødsling i slutten av vekstsesongen minsker plantenes evne til å bli godt herdet. Blir det frost for tidlig om høsten felles ikke bladene på vanlig vis. Avkastninglaget i enden av bladstilken dannes når dagene blir kortere og temperaturen synker. Hos f.eks. vintereik (Quercus petraea) blir bladfellingslaget dannet først om våren og eika beholder brune blad gjennom vinteren. Hvilken fordel de har av dette er uvisst. Frostakklimatisering og vinderforberedelser begynner på sensommeren. Fytokrom registrerer at daglengden endrer seg. Muligheten for frost varierer med breddegrad og høyde over havet slik at plantene må tilpasses voksestedet i form av økotyper eller provenienser. Fytokrom i planter i Nord-Norge må reagere på en annen daglengde enn fytokrom i en plante i Sør-Norge eller mellom-Europa. I bladhjørner hos busker og trær dannes det vinterknopper med anlegg for neste års skudd, blad og blomster. Første akklimatiseringsfase skyldes vekstavslutning og induseres av kortere dager, men også av vannstress. Lave temperaturer (2 - 5 oC) kan også medvirke. Hos noen busker og trær er ikke kort dag alene tilstrekkelig til å indusere vinterhvile, både kort dag og lav temperatur er nødvendig. Andre fase gir endringer av membranene og endringer i lipidsammensetningen i kloroplaster. En tredje fase kan induseres etter lengre eksponering ved ca.-10oC. Når vekstsesongen er slutt vil plantene kunne tåle gradvis lavere og lavere temperaturer. Akklimatiseringen utover høsten vil nå et platå hvor overlevelsestemperaturen er -5 - -10 oC. Etter en stund fortsetter akklimatiseringen til å tåle enda lavere temperatur.
Herdingsprosessen består av:
- 1) Bladene mottar signal om kortere dager som gir vekstavslutning.
- 2) Akkumulering av lagringsstoffer f.eks. stivelse og lipider. En nedgang i innholdet av fosfolipidet fosfatidylcholin under frysingen ser ut til å være korrelerte med frostskadene. Planter som er godt herdet beholder et høyt nivå av fosfatidylcholin. Mengden askorbat, arginin, prolin og redusert glutathion øker, og hos noen arter øker mengden sorbitol og mannitol. Økning i protein og RNA-konsentrasjon, mens DNA-innholdet er stabilt.
- 3) Temperatur fra 5oC og lavere starter det siste trinnet i herdingsprosessen..
- 4) Lav temperatur med påfølgende lange dager bryter hvilen om våren.
Det er en rekke miljøfaktorer som deltar ved induksjon og bryting av hvile: Daglengde, temperatur og næringstilgang. Økotypene har forskjellig krav til hvileinduksjon og hvilelengde. Sørlige økotyper har lang hvile sammenlignet med nordlige økotyper. I nordlige områder med lang og stabil vinter vil veksten hemmes av lav temperatur og det blir liten seleksjon for lang hvile. Lav temperatur bryter hvilen.

Vann utvider seg når det fryser. Om vinteren fryser vannet i apoplasten (cellevegger, intercellularrom og xylem), men vannet i protoplasten er underkjølt. Utvidelen kan få greiner og stamme på varmekjære løvtrær til å sprekke. Skaden blir dekket til av kallusvev.
Underkjølt vann i frostherdete urter, busker og trær
"Tidligsådd åker bør ingen tro". Fra Håvamål
Når det dannes is går vannmolekylene fra en tilfeldig plassering til et mer ordnet system. Mange væsker inkludert vann behøver ikke å fryse ved fryse- og smeltepunktet, men kan underkjøles. Tilføres det noe som kan tjene som utgangspunkt for iskrystalldannelse fryser løsningen raskt. Iskjerner katalyserer overgangen fra vann til is. Hvis en væske kontinuerlig mister varme vil temperaturen gå under frysepunktet til underkjølingspunktet. Straks isen fryser ved iskjernetemperaturen vil varmen fra krystalliseringen frigis og temperaturen stiger til en maksimal verdi (eksoterm topp). I en ren vannløsning vil eksoterm topp og frysepunktet være like. Homogen iskjernedannelse betyr at det dannes iskjerner spontant uten tilstedeværelse av fremmedlegemer i væsken. Rene vanndråper kan underkjøles til minus 38.1oC. Plantecellenes innhold av forskjellige stoffer senker ytterligere temperaturen for iskjernedannelse. Planteceller kan underkjøles til -41oC - -47oC avhengig av frysepunktdepresjon som kommer i tillegg.
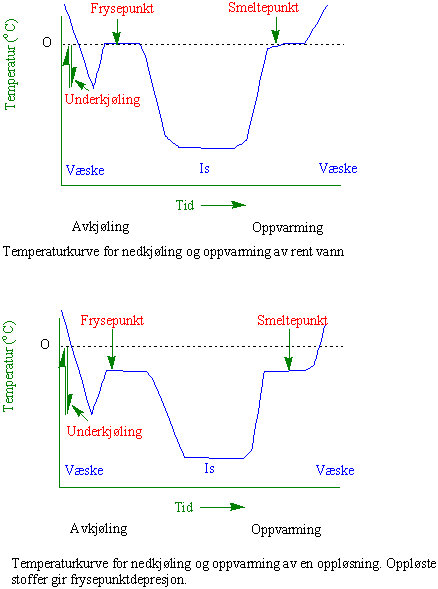
Heterogen iskjernedannelse er ikke-vann katalysert isdannelsen, dvs. isen starter fra fremmedlegemer i vannet. Antifrysproteiner hindrer iskrystaller å vokse. Frostherdige trær og busker lager is i cellevegger og intercellularrom og det må finnes iskjernedannere i disse områdene. I starten trengs det få iskjerner siden disse etterhvert gir sekundær iskjernedannelse. Antifryseproteiner som senker frysepunktet i oppløsninger er i planten lokalisert til apoplasten og kan modifisere iskrystallenes dannelse og form. Hvis en underkjølt væske tilsettes en iskrystall vil den fryse umiddelbart, men dette skjer ikke hvis den inneholder antifryseproteiner.
Iskjernedannende protein har også blitt funnet i bakterier som lever på bladoverflater. Pseudomonas syringae, som først ble funnet på overflaten av syrinblad (Syringa vulgaris), har vist seg å være spesielt effektiv til å lage proteinkjerner som danner is allerede like under frysepunktet for vann. Arter av Erwinia og Xanthomonas kan også dette. Iskjernedannende bakterier inneholder et ing gen(“ice nucleating gene”). Bakteriene har en fordel av at det dannes iskrystaller som ødelegger plantevevet slik at bakterien får tilgang til næringsstoffene i planten. F.eks. vil materiale fra Pseudomonas syringae fryse ved ca. -3.5 oC, mens et tilsvarende testoppsett uten bakterier fryser ved -17 oC. Iskjernedannere fra bakterier kan også brukes i snøkanoner. Genmodifiserte bakterier hvor det iskjernedannende genet ing var fjernet har blitt undersøkt om disse kunne gi økt frostresistens hos planter.
Både cellevegg og plasmamembran danner en effektiv barriere mot ekstracellulær is og sørger for at det blir en vannpotensialgradient mellom ut- og innside av cellen.
"Ja visst gör det ondt när knoppar brister. Varför skulle annars våren tveka ?" Karin Boye.
| Temperatur (oC) | is (e0, kPa) | Underkjølt vann (eo, kPa |
| 0 | 0.611 | 0.611 |
| -5 | 0.402 | 0.422 |
| -10 | 0.260 | 0.286 |
| -20 | 0.10 | 0.13 |
Metningsvanndamptrykket (eo) i kilopascal (kPa) over is og over underkjølt vann ved forskjellige temperaturer.Vanndamptrykket er lavere over is enn over vann ved samme temperatur. Det betyr at hardet først blitt dannet iskrystaller vil vann bevege seg fra underkjølt vann i omgivelsene til iskrystallene. Dannes det iskrystaller i apoplasten vil vann bevege seg ut av symplasten og over til apoplast.
Membraner og lav temperatur
Membranene fungerer som en barriere rundt kompartementer i cellen.Fettmolekylene i membranene danner et dobbeltlag hvor det er innleiret eller festet steroler og proteiner. Proteinene er enten perifere, som lett kan fjernes fra overflaten, eller integrerte proteiner som er en del av membranen. Proteiner kan være festet til fettsyrer med amidbinding til aminosyren glycin, eller festet til isoprener som farnesyl (C15) og geranylgeranyl (C20). Sterolene f.eks. sitosterol, camposterol og stigmasterol har en fri hydroksylgruppe og er med å stabilisere membranene. Kuldeaklimatisering består i å gjøre membranene kuldetolerante. Plasmamembranproteiner er festet til veggen med trådlignende strukturer kalt Hechttråder. Oppkalt etter Kurt Hect som oppdaget disse på begynnelsen av 1900-tallet. Hechttrådene påvirkes av lav temperatur. Kuldeakklimatisering gjør at mengden fosfolipider i membranen øker, mens mengden glukocerebrosider minsker. Glukocerebrosider er lipider hvor en av fettsyrene er sphingosin. Fettsammensetningen i thylakoidene endrer seg med temperaturen dvs. minsket forhold mellom galaktolipidene MGDG (monogalaktosyl diacylglycerol)/DGDG (digalaktosyl diacylglycerol) og økt innhold av umettet fettsyrer ved lavere temperatur.
Isolerende snøleier
Episoder med frost skyldes kald polarluft, eller stort varmetap fra jorda spesielt skyefrie netter vår og høst med stor utstårling fra jorda. I høyere alpine strøk kan frost forekomme hele året. Fjellplanter er bedre tilpasset til sommerfrost enn lavlandsplantene. Energibalansen på jorda er forskjellen mellom innkommet kortbølget stråling og utsendelse av langbølget stråling. Strålingsbalansen påvirkes av vinkel for innkommen stråling avhengig av breddegrad, årstid og helning på terrenget. Tografien som kan gi lav temperatur som følge av kalde luftstrømmer f.eks. i dalbunner. Barmarksfrost minsker muligheten for overlevelse av underjordiske organer. Dessuten hindres vann og næringsopptak i plantene når jorda er frosset. Frost trenger sakte ned i jorda. Det betyr at lav temperatur over lenger tid er nødvendig for at jorda skal fryse. Tørr jord fryser raskere enn våt jord. Vannet i jorda kan også underkjøles. Underkjølingen kan vare i flere timer og når vannet fryser frigis krystallisa sjonsvarme. Jorda holder 0oC så lenge som frysingen pågår. I tørr grovkornet jord trenger frosten ned med ca. 2 cm per dag. I våt humus jord ca. 0.6 cm per dag. Jordstrukturen kan endres ved frost f.eks. jordflyt (solifluksjon) og polygonmark i fjellet, . Iskrystalldannelse i jorda kan være med på å eksponere røtter ved frostheving og oppfrysing, hvor is hever det øvre jordlaget. Det er et stort hopp i temperaturgradienten mot jorda hvis varmetapet fra jorda reduseres med et isolerende lag med strøfall, mose eller snø. Snødekket begrenser hvor langt ned frosten går fordi snøen har isolerende luftlag og er en dårlig varmeleder. Forandringen i temperatur i takt med luftas temperatursvingninger skjer derfor på snøoverflaten og temperaturendringen transporteres sakte til de lavere deler av snølaget. Snøleier i fjellet gir beskyttelse mot lav temperatur, men ved sein avsmelting kan vekstsesongen reduseres betydelig og gi redusert mulighet for overlevelse. Fjellplanter kan starte med fotosyntese under snøen. De laveste temperaturene er fra 0-10 cm under jordoverflaten. Kommer snøen før jorda er frosset kan et 2 m tykt snølag holde temperaturen ved jordoverflaten ved 0 - -2oC. Et dekke av is eller kompakt snø kan imidlertidig også være skadelig. Diffusjonen av gass gjennom islaget går sent og konsentrasjonen av karbondioksid kan øke til opptil 45 % (v/v) og oksygenkonsentrasjonen kan synke til 3 % (v/v). Etanolproduksjon og Pasteur-effekter kan oppstå i plantevevet. Isbrann oppstår når sola skinner gjennom et isdekke ned på plantene under. Andre snøskader på trær og busker er bøyning, toppbrekk, splitting og velting. Trykket av snøen er proporsjonalt med snømengden og tettheten av snøen.
Skoggrense og temperatur
Lav vintertemperatur begrenser utbredelsen av mange plantearter. Frostskader er et viktig selektivt press i evolusjonen av mange planter. Skoggrensen blir lavere lenger mot nord, og når til slutt havnivået (Alexander von Humbolts lov). Ved ekvator er skoggrensen ca. 3.500- 4.000 m. Vi har en 1) klimatisk skoggrense som er den høyeste lokalitet der det vokser skog og som er begrenset av høyden over havet og ikke lokalklimatiske forhold og 2) en empirisk skoggrense som viser hvordan skoggrensen framtrer på et topografisk kart hvor klima, frømodning, vind, og snøskred er faktorer som innvirker. Det er altså en lineær sammenheng mellom skoggrense og breddegrad. Skoggrensen stiger i fjellstrøkene mot sør, men faller mot kysten og mot nord. I Norge lager bjerk den klimatiske skoggrense etterfulgt av furu og gran. Middeltemperaturen i de varmeste sommermånedene er også med å bestemmer skoggrensen. Hva er den fysiologiske mekanismen bak dette ? Trær trenger en minimum sommervarme tilsvarende 1 måned ved 10 oC. Frøproduksjonen hos trær skjer med irregulære intervaller f.eks. gran hvert 4-5 år. Ved større høyder over havet er det frøproduksjon kanskje bare hvert 8-11 år.
"Op i de bratte fjælde der enerbusken er kan ingen andre følge av alle skogens trær. Knut Hamsun "Enerbusken"
Naturkalender
Hvordan veksten av de forskjellige plantene preges av årstidene og været kalles fenologi. I begynnelsen av april vil hestehov (Tussilago farfara)og hassel (Corylus avellana) blomstre på Østlandet mens snøflekkene enda ligger. Deretter kommer or (Alnus), selje (Salix) og hvitveis (Anemone nemorosa) og i begynnelsen av mai starter våronna. I midten av mai starter blomstringen av marianøklebånd (Primula veris) og liljekonvall (Convallaria majalis). I slutten av måneden er hegg (Prunus padus) og løvetannen (Taraxacum) i fullt flor og det er tid for hageeieren å så bønner og mer varmekrevende frø. Så kommer plantene på rekke og rad med blomster utover sommeren. I begynnelsen av august begynner røsslyng (Calluna vulgaris) å blomstre og bikubene settes ut i marka for at biene skal samle lynghonning. I begynnelsen av oktober starter trærne med å bli gule og felle løvet. En slik utvikling gjennom årstidene er koblet til når forskjellige trekkfugler kommer og reiser, og når det er på tide og høste og så. Hvert sted i Norge har sin egen naturkalender.
Høstfarger
Hos flerårige løvtrær og busker skjer det om høsten en nedbrytning av klorofyll og proteiner i fotosynteseapparatetsom. I klorofyllnedbrytningen fjernes først fytolhalen og klorofyllmolekylet omdannes til klorofyllid ved hjelp av enzymet klorofyllase. Deretter fjernes magnesium (Mg2+ ) i chelat i tetrapyrollringen katalysert av dechelatase og det dannes feoforbid. Neste trinn er en oksidering som gir fluorescerende klorofyllnedbrytningsprodukter katalysert av feoforbid oksygenase. En hypotese som forklarer hvorfor klorofyll må nedbrytes før karotenoidene går ut på at klorofyll og sannsynligvis porfyrin må fjernes fordi de kan gi toksiske fotodynamiske effekter forårsaket av singlett oksygen. Gulfargede karotenoider beholdes så lenge som mulig for å hindre oksidative skader. Nitrogen i klorofyll utgjør bare 3 - 5 % av totalt nitrogen i bladet og betyr lite i det totale N-budsjett, sammenlignet med det nitrogen som befinner seg i proteiner. Karotenoidene inneholder ikke nitrogen og det kan være en grunn til at de ikke tas vare på. Innholdet av proteaser i bladet inkludert cystein protease øker utover høsten. Det skjer en kontrollert celledød i avkastingslaget mellom bladstilk og grein. Aldringsrelaterte gener skrus på . Hvis nettene er kalde og dagene solrike lages det mye rødfargete anthocyaniner. Planter som beholder bladene gjennom vinteren har glinsende læraktige blad tilpasset vintertørke.