Ornithin-syklus, arginin, glutaminsyre og prolin
Glutaminsyre (glutamat) spiller en sentral rolle i assimilasjonen av nitrogen inn i organiske forbindelser i form av ammonium. Glutaminsyre ble første gang isolert av Ritthausen i 1866 ved hydrolyse av gluten fra hvete og derav navnet. Glutamin er amidet av glutaminsyre. Glutaminsyre er råmaterialet for andre aminosyrer ved å bidra med karbonskjeletter og nitrogen. Arginin, glutamin og asparagin er viktige for lagring og transport av nitrogen i planter. Arginin ble første gang oppdaget av Schulze og Steiger i 1886 og isolert fra kimplanter av lupiner. Nitrogenet frigis fra arginin i form av urea katalysert av enzymet arginase, og produktet er ornithin. Citrullin er en aminosyre som man fant store mengder av i saften av vannmelon (Citrullus vulgaris). Det skulle vise seg at citrullin var et mellomprodukt i biosyntesen av arginin i ornithin-syklus (Krebs-Henseleit syklus) oppdaget i 1932. Ornithin reagerer med karbamoylfosfat og gir citrullin. Citrullin omdannes deretter via argininosuccinat til arginin. Ornithin lages forøvrig fra glutamat via metabolismerekken glutamat, acetyl-glutamat, acetyl-ornithin og ornithin.I tillegg er arginin utgangsmateriale for å lage signalstoffet og radikalet nitrogenoksid (NO) katalysert av enzymet NO syntase. Arginin inngår også i opiner (octopin) laget av bakterien Agrobacterium tumefasciens som gir krongalle på planter.
{kind=link}
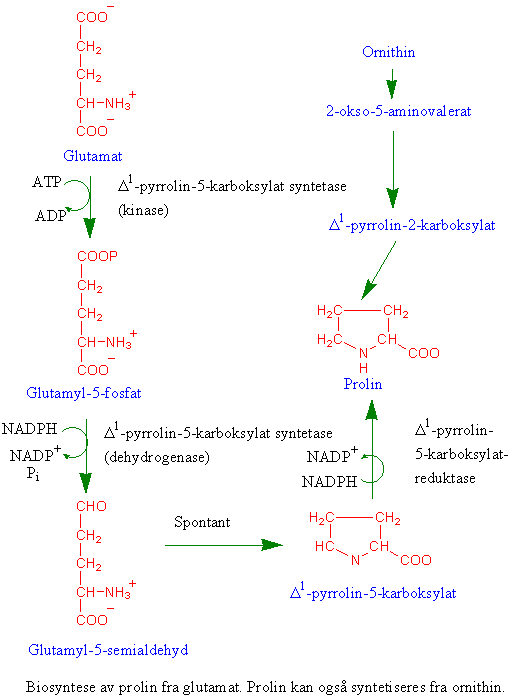
Stressaminosyren prolin
Prolin lages fra aminosyren glutamat som omdannes først til glutamat semialdehyd som videre gir D-pyrrolin-5-karboksylat katalysert av D-pyrrolin-5-karboksylat syntetase. D- pyrrolin-5-karboksylat reduseres til prolin av D-pyrrolin-5-karboksylat reduktase. Prolin akkumuleres under forskjellige typer stress f.eks. saltstress. Plantehormonet abscisinsyre deltar i aktiveringen av D-pyrrolin-5-karboksylat syntase. Celleveggene inneholder protein med mye hydroksyprolin. Hvis det er stort behov for prolin til celleveggoppbygging kan argininkatabolisme bidra med prolin.
{kind=link}
Asparagin
Vauquelin og Robiquet fant i 1806 den første aminosyren som var asparagin og denne ble isolert fra asparges. Amidnitrogenet i glutamin kan overføres til aspartat katalysert av asparagin syntetase:
\(\displaystyle aspartat + glutamin\; _{\overrightarrow {asparagin\; syntetase}}\; asparagin + glutamat + AMP + PP_i\)
Enzymet finnes i cytosol i røtter og blad og i nitrogenfikserende rotnoduler. I mørke, hvor det blir lite karbonskjeletter i forhold til nitrogen, lages asparagin på bekostning av glutamin. Lys, som aktiverer glutamin syntease, hemmer både aktivitet og genuttrykk av asparagin syntetase.
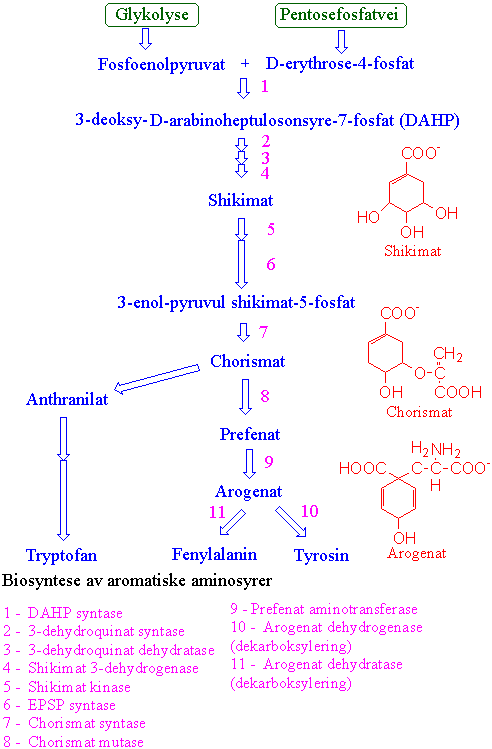
Shikimatbiosyntesevei og syntese av aromatiske aminosyrer
Shikimatbiosynteseveien finnes i planter, bakterier og sopp, men ikke hos dyr. Shikimatbiosynteseveien finnes i planter, bakterier og sopp, men ikke hos dyr. Den har fått navn etter det japanske navnet shikimino-ki for planten kinesisk stjerneanis (Illicium religiosum, Illiciaceae). Biosynteseveien fører fram til de 3 aromatiske aminosyrene fenylalanin, tyrosin og tryptofan. Metabolismeveien starter med erythrose-4-fosfat som kommer fra pentosefosfatveien, og fosfoenolpyruvat fra glykolysen. Det første produktet er 3-deoksy-D-arabinoheptulosonsyre- 7-fosfat (DAHP) katalysert av 3-deoksy-D-arabino-heptulosonat-7-fosfat syntase. Dette første enzymet blir ikke hemmet av noen av de aromatiske aminosyrene. Deretter følger en defosforylering, tilføring av vann og reduksjon med NADPH som gir shikimat. Shikimat fosforyleres med ATP og reagerer med fosfoenolpyruvat og danner 3-enolpyruvylshikimat-5- fosfat (EPSP).Enzymet 5-enolpyruvul-shikimat-3-fosfat syntase (EPSPS) katalyserer overføring av enolpyruvyl delen av fosfoenolpyruvat til shikimat-3-fosfat hvor man får dannet chorismat. Dette enzymet hemmes av ugrasmiddelet glyfosat. Chorismat er greiningspunkt og siste felles forbindelse mellom tryptofan og fenylalanin/tyrosin. Chorismat omdannes til prefenat, katalysert av chorismat mutase, som videre ved transaminering gir arogenat. Arogenat gir tyrosin etter dekarboksylering katalysert av arogenat dehydrogenase. Arogenat kan også ved dekarboksylering og fjerning av vann gi fenylalanin, katalysert av arogenat dehydratase. Det finnes to isoenzymer av chorismat mutase. En type hemmes av tyrosin og fenylalanin, og aktiveres av tryptofan. Det andre blir ikke regulert ved tilbakekobling. Arogenat dehydratase hemmes av fenylalanin og aktiveres av tyrosin. Arogenat dehydrogenase hemmes av tyrosin. I biosynteseveien til tryptofan med en indolring omdannes chorismat til anthranilat katalysert av anthranilat syntase, med glutamin som aminodonor. Deretter følger 5 enzymatiske trinn hvor indolringen lages. Tryptofan benyttes også til å lage auxin, alkaloider og fytoaleksiner. Alle trinnene i syntesen av aromatiske aminosyrer skjer i plastidene, men isoenzymer i synteseveien finnes også i cytosol. Biosynteseveien kan aktiveres ved stresstilstander.
{kind=link}
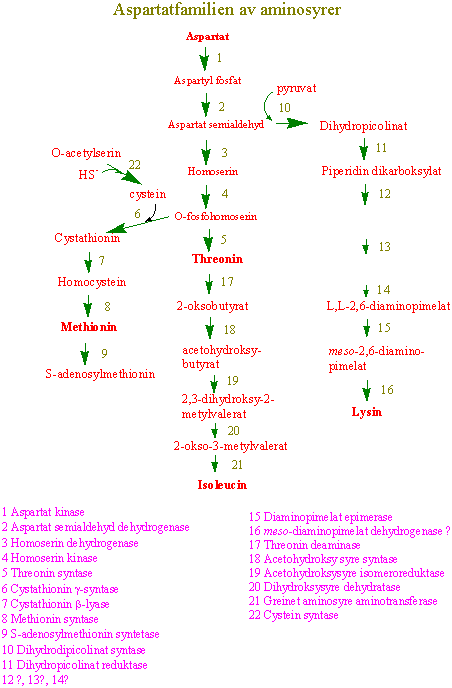
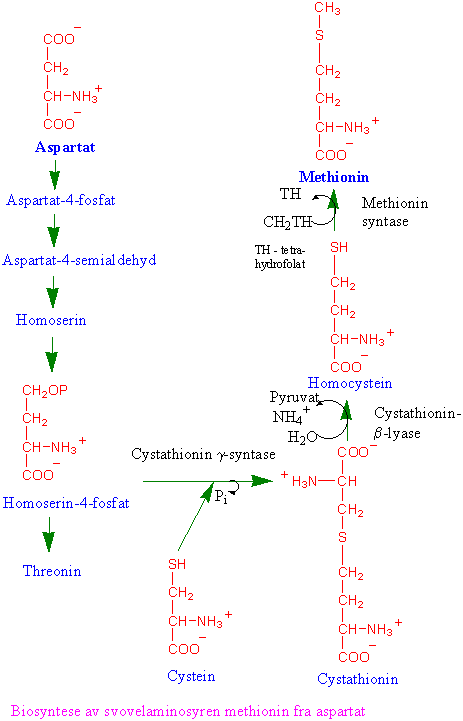
Aspartatfamilien av aminosyrer
Lysin, threonin, methionin, de greinete aminosyrene leucin, isoleucin og valin; og de aromatiske aminosyrene fenylalanin og tryptofan hører med til de 7 såkalte essensielle eller nødvendige aminosyrene i kostholdet for mennesker. Vi får tilført aminosyrene via proteiner i dietten. Det er bare planter og mikroorganismer som klarer å lage disse livsviktige aminosyrene. Ikke alle proteiner inneholder en balansert fordeling av disse aminosyrene tilpasset vår ernæring. Spesielt protein fra korn inneholder lite lysin, slik at en ensidig diett basert på kornprodukter uten annen proteintilførsel kan gi for lite lysin. Dette kan oppveies noe ved samtidig å spise protein fra erteplanter. Barn, spesielt utenom industrilandene, kan få for lite av de nødvendige aminosyrene og det kan resultere bl.a. i sykdommen kwashiorkor, som på afrikansk betyr "tatt for tidlig fra brystet". Sykdommen gir oppsvulming av magen og dårlig utvikling av hjerne og muskelceller. Lysin, threonin, isoleucin og methionin lages i en greinet biosyntesevei med asparaginsyre som utgangsmateriale - aspartatfamilien av aminosyrer. Det første enzymet i biosynteseveien, aspartat kinase, og enzymene i greiningspunktene er under streng metabolsk kontroll av endeproduktene. En overskrift i Aftenposten 13.8.1979 var "Produksjon av barnegrøt med lysin opphørt" etter at det skapte furore da det ble kjent at lysin ble tilsatt barnematen. Slik tilsetning er imidlertid vanlig i dyrefor.
{kind=link}
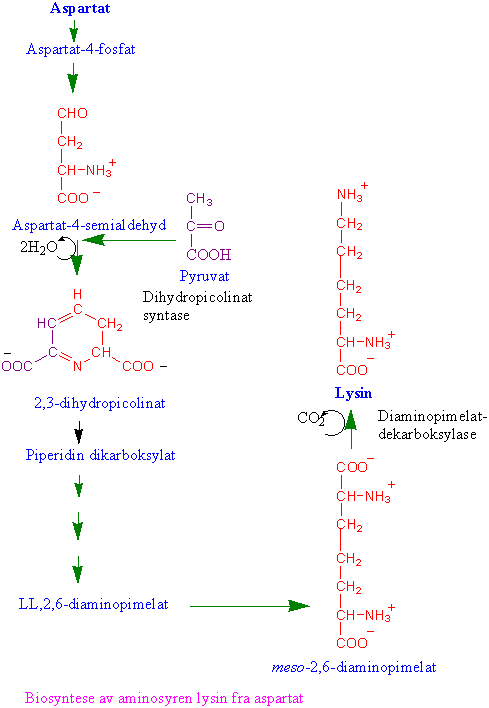
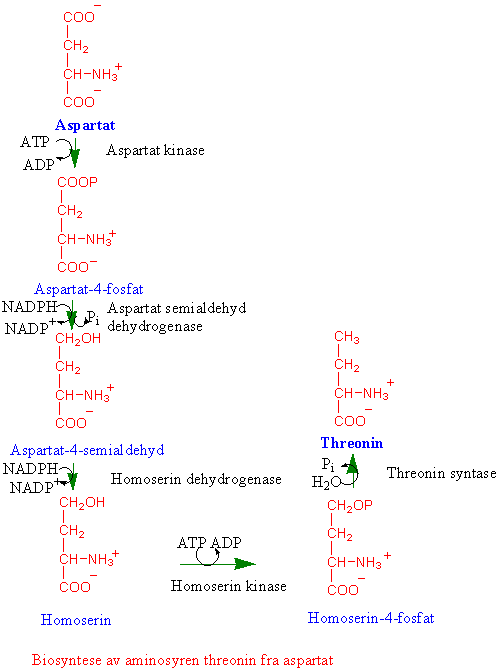
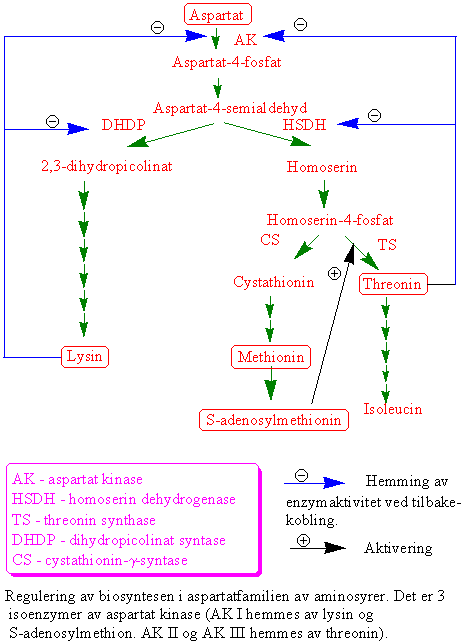
Asparaginsyre omsettes i en greinet biosyntesevei til aminosyrene lysin, methionin og threonin (aspartatfamilien av aminosyrer). Reguleringsenzymer i biosynteseveien er aspartat kinase som fosforylerer aspartat og gir aspartat-4-fosfat og feedback reguleres av lysin og threonin. Homoserin dehydrogenase reguleres av threonin; og dihydropicolinat syntase på veien til lysin som kondenserer 3-aspartatsemialdehyd og pyruvat og gir 2,3-dihydropicolinat som hemmes av lysin. Biosyntesen av isoleucin og valin har flere felles enzymer, og står i nær tilknytning til biosyntesen av leucin. Threonin er utgangsmateriale for å lage isoleucin. Methionin kan omdannes til S-adenosylmethionin. S-adenosylmethionin er en viktig metyldonor i metyleringsreaksjoner og er forkomponent til etylen og polyaminer. Svovelgruppen i cystein overføres til methionin via homoserin-4-fosfat, cystathionin og homocystein. Metylering av homocystein skjer ved hjelp av tetrahydrofolat. Ved spiring av erter dannes det store mengder homoserin. Det finnes 3 forskjellige isoenzymer, AKI, AKII og AKIII. AKI hemmes av lysin og S-adenosylmethionin. Både AKII og AKIII hemmes av threonin. Hvis man lar embryoer av bygge vokse i nærvær av lysin og threonin som hemmer AK vokser de dårlig siden det blir for lite methionin. Dihydrodipicolinat syntase, som er det første enzymet i greiningspunktet til lysin, blir hemmet av lysin ved tilbakekobling. Homoserin dehydrogenase (HSDH), som reduserer aspartat-4-semialdehyd til homoserin reguleres av threonin og og danner et stort enzymkompleks med threoninfølsom aspartat kinase. Hele biosynteseveien finnes i plastidene, bortsett fra siste trinnet i syntesen av methionin og S-adenosylmethionin som finnes i cytoplasma.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
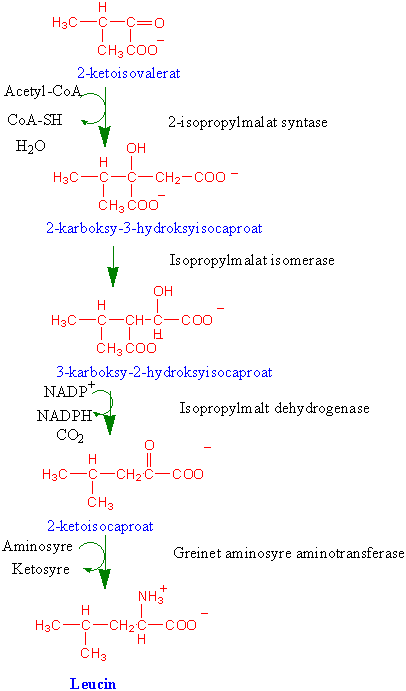
Biosyntese av greinete aminosyrer (isoleucin, valin og leucin)
Første trinnet fra threonin til isoleucin katalyseres av anabolsk threonin deaminase. Threonin deaminase hemmes av isoleucin ved tilbakekobling. Det finnes threonin deaminase som deltar i katabolisme av threonin og serin, men dette enzymet hemmes ikke av isoleucin. Neste trinn er overføring av 2 karbonatomer fra hydroksyetylpyrofosfat katalysert av acetohydroksysyre syntase (acetolaktat syntase). Dette enzymet hemmes av herbicider (ugrasdrepere) av typen sulfonylureaforbindelser, triazolpyrimidiner og imidazolinoner. Syntesen av valin skjer parallelt med isoleucin, og disse to biosynteseveiene har fire felles enzymer. Første trinnet i syntesen av leucin katalyseres av isopropylmalat syntase, som hemmes av leucin ved tilbakekoblingen. I dette trinnet overføres acetyl via acetyl-CoA til 2- ketoisovalerat.
Fra fenylalanin til fenoler og flavonoider
Fenylalanin danner utgangspunkt for fenoler. Første trinn katalyseres av fenylalanin ammonium lyase (FAL) som fjerner ammonium og danner trans-cinnamat. Aktiviteten til FAL øker ved mangel på nitrogen eller fosfor, men også ved generell dårlig næringstilgang. Aktiviteten til FAL øker også i lys og ved lav temperatur, og kan også påvirkes av soppinfeksjon. Reguleringen av FAL skjer på transkripsjonsnivå. Rødfarede anthocyaniner som kommer fra fenylalanin dannes på solsiden av et eple. Høstblader kan danne mye rødfarge (anthocyaniner) hvis høsten er solrik og kald. Dette er eksempler på lysregulering av FAL. Cinnamat omdannes til para-coumarat, en C6-C3-forbindelse, ved tilførsel av en hydroksylgruppe i 4-stilling katalysert av cinnnamat 4-hydroksylase, - en cytokrom P450 monooksygenase.
para(4)-coumarat brukes til å lage caffeinsyre, coumariner og er råmateriale for syntese av lignin. para-coumarat kan reagere med CoA og danne 4-coumaroyl-CoA katalysert av 4- coumarat:CoA ligase:
Chalkon syntase kobler sammen 3 molekyler malonyl-CoA og para-coumaryl-CoA og danner chalkon. Chalkon kan omdannes til flavoner, isoflavoner (isoflavonoider) og dihydroflavonoler. Dihydroflavnoler gir igjen utgangspunkt for anthocyaniner og kondenserte garvestoffer (tanniner).
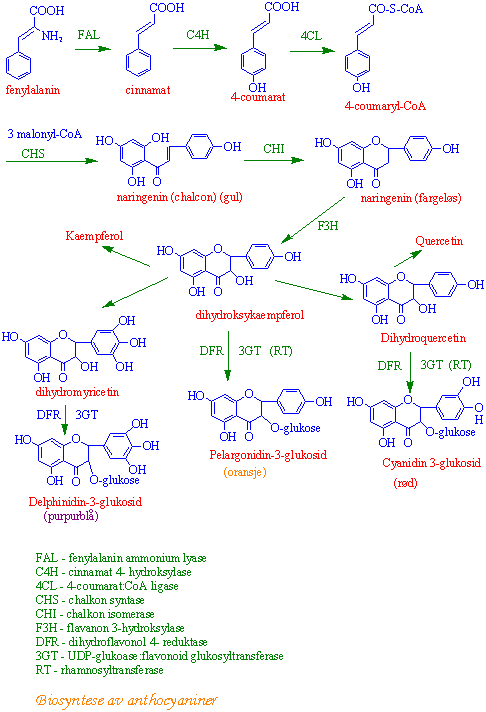{kind=link}
Flavonoider fra shikimatveien
Flavonoider er en heterogen gruppe fenolforbindelser bestående av 15 karbonatomer koblet sammen som C6-C3-C6. De er satt sammen av to ringer A og B, hvor B-ringen er C6-C3 som kommer fra shikimatveien. para-coumaryl-CoA reagerer med 3 molekyler malonyl-CoA katalysert av chalcon syntase. Chalkon syntase er et viktig reguleringsenzym. Flavonoidene klassifiseres etter oksidasjonstrinnet på C3 broen mellom de to C6-ringene.
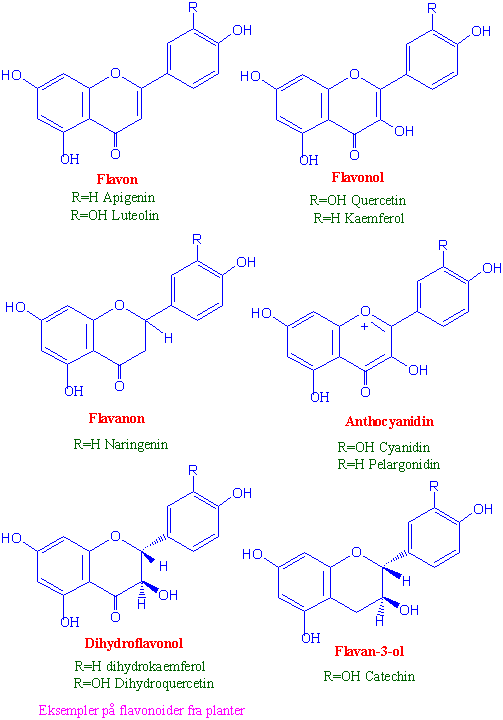{kind=link}
Lignin
Lignin er en polymer i sekundærveggen hos planter som gir mekanisk støtte i hudvev og i vanntransportsystemet i planter. Lignin gir også beskyttelse mot sykdomsfremkallende sopp, bakterier og virus. Lignin virker som "betong" i celleveggen hvor cellulose og hemicellulose danner "armeringsjernet". Lignin er et stort og meget komplekst makromolekyl bygget opp av 3 forskjellige fenylpropanoider (C6-C3): coniferylalkohol, sinapylalkohol og 4-coumarylalkohol. Alle disse 3 er vanlig i lignin fra angiospermer. Lignin fra gymnospermer inneholder mest coniferylalkohol og mindre mengder 4-coumarylalkohol. Hydrogenperoksid og peroksidaser deltar i en fri radikal polymerisering av fenylpropanoidene. De fleste organismer, untatt noen hvitråtesopp, har vanskeligheter med å nedbryte lignin. Lignin som isoleres fra planter er brunfarget og uløselig i vann. Lignin kan brukes til produksjon av syntetisk vanilje (vanillin).
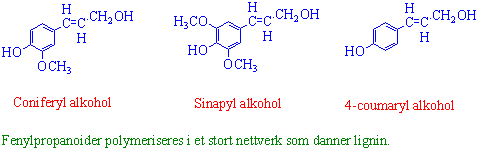{kind=link}
Lignaner
Lignaner er fenylpropanoiddimere som er bundet sammen i de alifatiske C3-sidekjedene f.eks. pinoresinol fra harpiks hos gran og furu. Lignaner er fargeløse og finnes i tillegg i kjerneved og blader. Noen av dem er virker som antioksidanter. Andre eksempler på lignaner er sesamin (Sesamum sp.), podophyllotoksin (Podophyllum sp.), og cubebin i frukten fra Piper cubeba. Olivil finnes i harpiksen som skilles ut av oliventrær (Olea europaea) og guaiaretinretinsyre i harpiks fra Guajacum officinale. Det finnes forskjellige hovedtyper lignaner.
Isoprenoider: to synteseveier fram til femkarbonforbindelsen isopentenyl difosfat
1) Mevalonsyrebiosynteseveien finnes i cytoplasma. Glukose omdannes via triosefosfat til acetyl-CoA som er utgangsmateriale. I denne biosynteseveien lages mevalonsyre (mevalonat) fra 3 molekyler med acetyl-CoA. Først lages acetoacetyl-CoA fra 2 molekyler acetyl-CoA katalysert av acetylCoA acetyltransferase. Acetoacetyl-CoA reagerer med ytterligere ett molekyl acetyl-CoA og danner b-hydroksy-b- metyl-glutaryl CoA (HMG-CoA) katalysert av HMG-CoA syntase. HMG-CoA reduseres deretter til mevalonsyre med NADPH som elektrondonor katalysert av HMG-CoA reduktase. Deretter følger to fosforyleringer med ATP (mvalonsyre kinase og fosfomvalonat kinase) som gir mevalonsyre-5-pyrofosfat (MVA-PP). MVA-PP dekarboksyleres ved hjelp av ATP (disfosfomevalonat dekarboksylase) og danner isopentenyl-pyrofosfat (IPP). IPP isomerase danner ved isomerisering dimetylallyl-pyrofosfat (DMAPP). IPP og DMAPP er de to femkarbonforbindelsene som danner grunnlaget for syntese
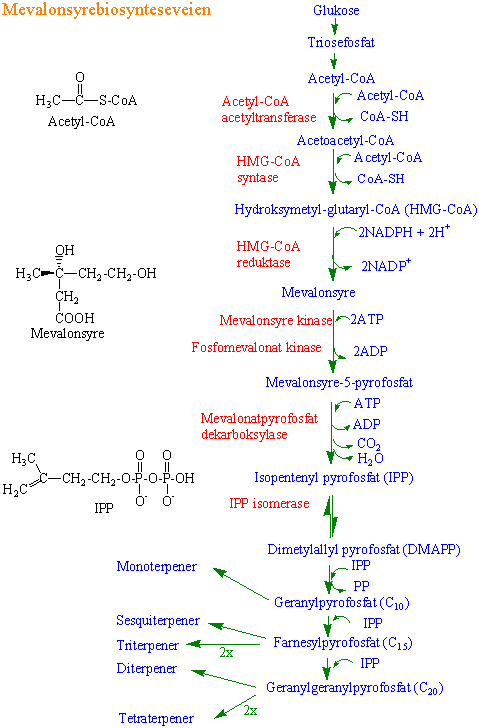{kind=link}
av isoprenforbindelser (isoprenoider).
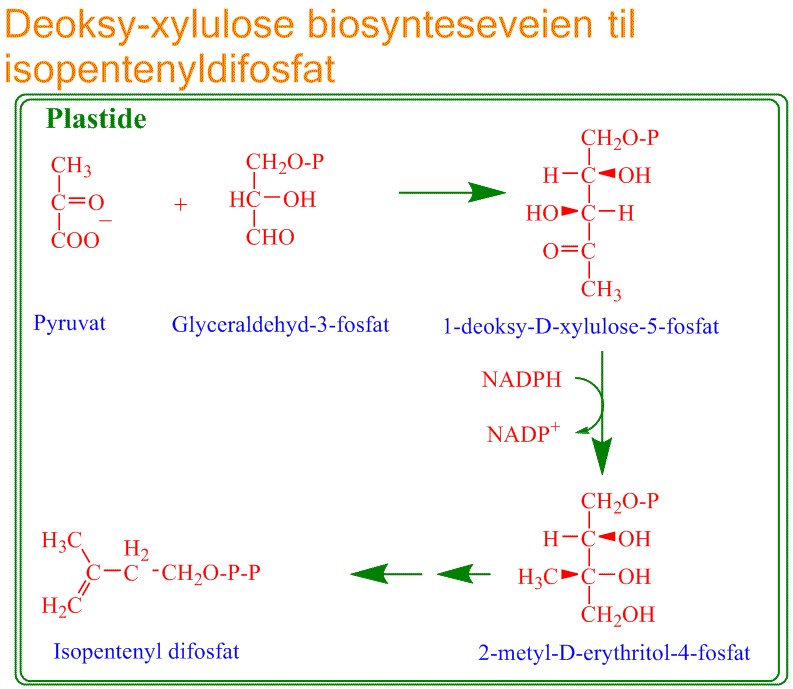
2) Deoksyxylulosefosfatbiosynteseveien finnes i plastider.
Glyceraldehyd-3-fosfat som kommer fra glukose danner sammen med pyruvat forbindelsen 1-deoksyxylulose-5-fosfat (DOXP) katalysert av DOXP syntase. DOXP omdannes via 2-metyl-D- erythritol-4-fosfat katalysert av DOXP reduktoisomerase til isopenteyldifosfat.
Monoterpener(C10), sequiterpener (C15), diterpener (C20) og polyterpener ([C5]n)
Alle terpener i planter settes sammen av 5-karbonenheter fra isopentenylpyrofosfat (IPP) og dimetylallyl pyrofosfat (DMAPP). 5-karbonenhetene blir ofte kalt isopren. Mange av terpenene virker som antibeitestoffer i harpiks. I tillegg har de funksjoner som plantehormoner (gibberelliner, abscisinsyre, brassinosteroider), pigmenter (karotenoider), membrankomponenter (steroler)
Enzymet isopentenyldifosfat isomerase katalyserer isomerisering mellom isopentenylpyrofosfat (IPP) og dimetylallyldifosfat (DMAPP). Limonen syntase kobler sammen IPP og DMAPP og lager geranyldifosfat. Geranyldisofat brukes til å lage (-)-limonen katalysert av plastideenzyymet 4s- limonen syntase. (-)-limonen kan nå enten brukes til å lage (- )-carvon eller (-)-mentol. Limonen-6-hydroksylase lager trans- carveol fra limonen som brukes til å lage carvon. 4s-limonen- 3-hydroksylase lager trans-isopiperitenol som brukes til å lage mentol bl.a. i trichomer hos peppermynte.
IPP og DMAPP danner geranylpyrofosfat bestående av 10 karbonatomer, og som er utgangspunkt for syntese av monoterpener. Geranylpyrofosfat ragerer med IPP og danner C15- forbindelsen farnesylpyrofosfat som brukes til å lage sesquiterpener. Nok en reaksjon med IPP lager geranylgeranylpyrofosfat med 20 karbonatomer og utgangspunkt for diterpener. To molekyler farnesylpyrofosfat gir molekyler med 30 karbonatomer som squalen, triterpener og steroider. To molekyler geranylgeranylpyrofosfat kan brukes til å lage phytoen og tetraterpener med 40 karbonatomer.
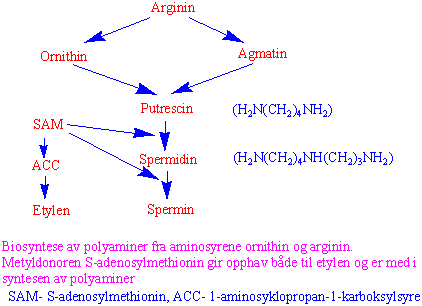
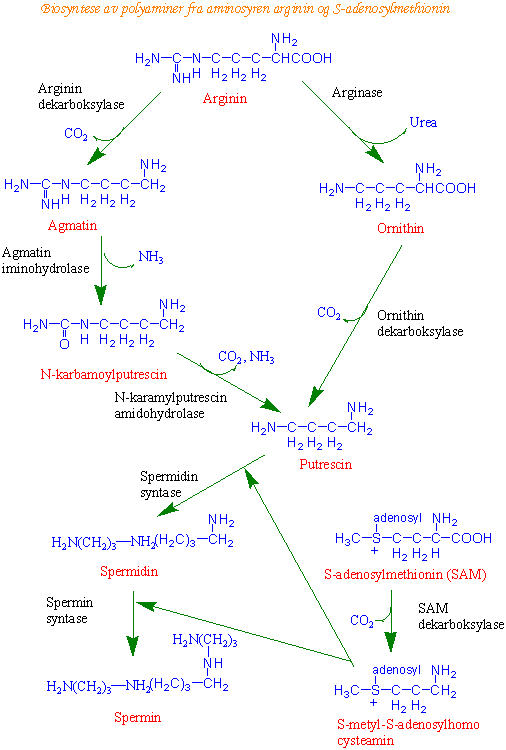
Biosyntese av polyaminer
Putrescin (et diamin) og polyaminene spermidin og spermin har viktige reguleringsfunksjoner i plantene. Både diaminer og polyaminer virker som organiske polykationer og kan binde til DNA, spesielt i plastider og mitokondrier hvor DNA ikke er stabilisert av histoner. Diaminet cadaverin som kommer fra dekarboksylering av aminosyren lysin, er funnet hos arter i erteblomstfamilien. Dekarboksylert S-adenosylmethionin virker som propylamindonor i biosyntesen av polyaminer. S-adenosylmethionin (SAM) dekarboksyleres og gir S-metyl-S-adenosylcysteamin katalysert av SAM dekarboksylase. Propylaminogruppen overføres via enzymet putrescin aminopropyltransferase. Aspartat b- semialdehyd kan også virke som propylaminodonor og kan være en alternativ syntesevei til spermidin og homospermidin.
{kind=link}
{kind=link}