- DNA-molekylet
- DNA i kromosomer
- B-kromosomer
- Generasjonsveksling
- Replikasjon - fra DNA til DNA
- Tandemrepetert DNA
- Kloroplastgenomet
- Mitokondriegenomet
- DNA innhold og C-verdiparadoks
- Kjernegenomet og kromosomer
- Kosuppresjon
DNA-molekylet
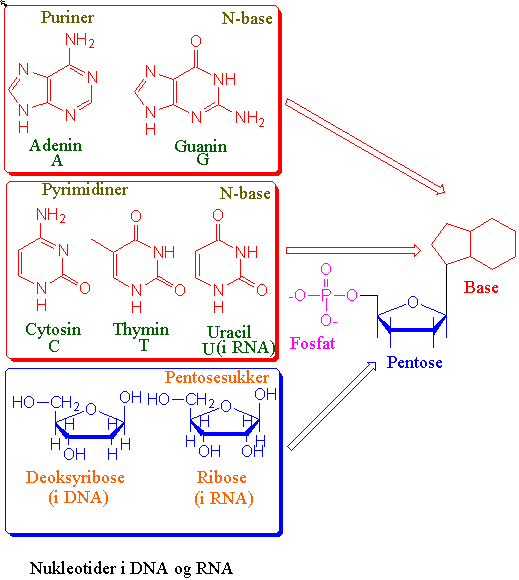
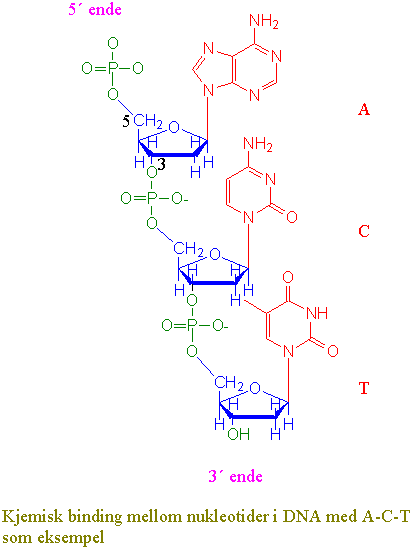
Den arvelige informasjonen ligger, untatt for noen RNA-virus, i DNA-molekylet. DNA overføres fra foreldre til avkom via spermier (pollen) og egg. DNA inneholder all informasjon som er nødvendig for å gjenskape et nytt individ. DNA (deoxyribonucleic acid) består av to komplementære tvinnete DNA-tråder som gir en høyredreiet dobbelheliks formet som en spiraltrapp. Hver DNA-tråd er en kjede med nukleotider. Hver nukleotid innholder en sukkerpentose (deoksyribose), en fosfatgruppe og en base. Basene kan være purinene adenin (A) og guanin (G), og pyrimidinene cytosin (C) og thymin (T). Sukkerfosfatene danner "gelenderet" og basene "stigtrinnene" i dobbelheliksen. Fosfat og sukker gir den ytre skjelettstrukturen. Sukker og fosfat bindes kovalent i en 5´-3´-fosfodiesterbinding. Strukturen 5´ til 3´ er et resultat av en fosfatester mellom 3´-hydroksylgruppen på sukker i den ene nukleotiden og 5´-fosfatgruppen i den neste nukleotiden. Den ene DNA-tråden vil derfor få en fri fosforylgruppe i 5´-enden og en fri hydroksylgruppe i 3´-enden. Ved konvensjon er DNA-sekvensen skrevet i den rekkefølgen den transkriberes dvs. fra 5´- til 3´- ende. De to DNA-trådene, motsatt rettet, er bundet sammen med hydrogenbindinger mellom basene. Adenin bindes til thymin (A-T) med 2 hydrogenbindinger. Guanin bindes til cytosin (G-C) med 3 hydrogenbindinger. De to enkelttrådene holdes godt sammen og gir en meget stabil struktur. Det er 10 baser for hver omdreining av heliksen. Når man skriver opp DNA sekvensen utelater man sukker og fosfat og angir bare basene. Basene A, T, G, og C angis som basepar (bp). DNA finnes i alle levende celler. Hos plantene finnes DNA i cellekjernen og i plastider og mitokondrier. Oppbygningen av DNA i disse organellene er forskjellig. Sekvensen av basene i DNA er karakteristisk og spesifikk for hver art.
{kind=link}
DNA i kromosomer
Arvematerialet DNA er pakket i kromosomer. Kromosomene i planter er bare synlig i et lysmikroskop i visse stadier av celledelingen, og er lettest å observere i mitosens metafase. Røntgendiffraksjon og elektronmikroskopi er nødvendig for å kunne observere finere strukturer i DNA.
De homologe kromosomene i hvert kromosompar har en karakteristisk struktur, og de fleste kromosomene kan identifiseres ut fra utseende og hvordan de farges. Kromosomene i cellekjernen inneholder en DNA-dobbelheliks hvis det ikke har skjedd replikasjon. Lengden på DNA-molekylet er ofte flere ganger lenger enn diameteren til cellen. DNA kan brytes i biter, og kromosomfragmenter kan mistes under celledeling. Derfor er det viktig at DNA er godt pakket og omgitt av kromosomale proteiner kalt histoner. Disse proteinene består av 5 hovedklasser med histoner (H1, H2A, H2B, H3 og H4) og en heterogen blanding av ikke- histonproteiner, ofte kalt høy mobilitetgruppeproteiner (HMG), og noen er assosiert med RNA. Histonene har høy affinitet til hverandre og vil sammenbindes. Komplekset bestående av DNA og protein kalles kromatin. DNA og kromatinfibren er tvunnet og kveilet opp om selv selv. Kromatinfibrene er lokalisert til spesielle steder i kjernen. Den indre overflaten av kjernemembranen inneholder spesielle proteiner kalt laminer som danner et fibrøst nettverk hvor kromatinfibrene er bundet. Kromatinet er satt sammen i en primær struktur i form av nukleosomer. Videre en sekundærstruktur i form av spoler med 6-7 nukleosomer per omdreining, en tertiærstruktur i form av løkker, og en kvartærstruktur for en avsluttende foldingen av kromatinet i kromosomer. Hvert nukleosom består av en oktamer av 8 histonproteiner og rundt denne er det tvunnet ca. 200 basepar med DNA, som perler på en snor, hvor perlen er nukleosomet og tråden er DNA. Histonene inneholder en hydrofob kjerne og har høyt innhold av basiske aminosyrer som reagerer med det sure DNA.
Enden av kromosomet kalles telomerer (gr. telos - ende; meros - del), og blir kortere og kortere for hver celledeling. Telomerer er assosiert med heterokromatin og kan inneholde basesekvenser som f.eks. (TTTAGGG)n eller TT(T/A)GGG. Telomerenes oppgave er å beskytte de to endene i kromosomet, for å hindre at vitale deler i enden blir borte under hver celledeling. DNA polymerase binder seg til enden av DNA og vil blokkere for kopiering av den siste biten av tråden. Enzymet som sørger for replikering av telomerene kalles telomerase.
Nukleolusorganiserende region (NOR) i kromosomet inneholder repeterte sekvenser som koder for mutiple kopier av RNA gener. Kjerneorganiserende region er forbundet med visse deler av kjernen kalt nukleolus, og det er her ribosomalt RNA (rRNA) blir transkribert og sammenbundet.
Kromosomene er sammensnørt ved centromeren, som inneholder spesielle repeterte sekvenser og spesielle proteiner. Centromeren er stedet for atskillelse av søsterkromatider under celledelingen. Flytting av kromosomene skjer ved en reaksjon mellom kinetokoren, en sammensatt struktur tilkoblet centromeren, og mikrotubuli. Kromosomet består av to hovedklasser av DNA: eukromatin og heterokromatin. Det antas at eukromatin er mest aktivt. Mindre aktivt heterokromatin inneholder mest repeterte sekvenser og er mest kompakt.
Det går an å identifisere alle kromosomene i en organisme ut fra størrelse, korte og lange armer, og nukleolusorganiserende region (NOR), og in situ hybridisering. Et diagram av de karakteriske egenskapene til kromosomene kalles idiogram (gr. idios - bestemt; gramma - tegning). Kromosomstrukturen kan beskrives som metasentrisk, akrosentrisk eller telosentrisk, med angivelse av telomere i enden av kromosomet og avsnøring ved centromeren. Hele settet med kromosomer i en organisme kalles genomet. Posisjonen til et gen på kromosomet kalles locus. I organismer som reproduserer seg seksuelt kan kromosomene deles i 1) autosomer, som er felles for begge kjønn, og 2) kjønnskromosomer. Plantene mangler ofte egne kjønnskromosomer.
B-kromosomer
B kromosomer atskiller seg fra vanlige A kromosomer, finnes i over 1000 plantearter og kommer i tillegg til de vanlige kromosomene. B kromosomene er mindre enn A kromosomene, inneholder mer heterokromatin, inneholder ikke sentralt viktige gener, og de nedarves på ikke-mendelsk måte. Man antar at B kromosomene kommer fra trisomi autosomvarianter av A-kromosomer. B kromosomene inneholder homologe sekvenser til deler av A kromosomer. Antallet B kromosomer i ville plantearter kan være opptil 8, og overføringen under celledeling favoriseres hvis de er i par. Noen kromosomer hos planter har et ekstra segment på et bestemt kromosom. Disse ligner på B kromosomer og inneholder mye heterokromatin. Polytenkromosomer finnes hos planter, men er dårlig karakterisert.
Generasjonsveksling
En celle med ett sett kromosomer er haploid. Har den to sett kromosomer er den diploid. Den diploide frøplanten kalles sporofytt fordi den ved reduksjonsdeling lager to slags sporer: pollenkorn og embryosekk. De haploide "plantene" som lages ved utviklingen av pollenkorn og embryosekk kalles gametofytter, fordi de danner kjønnsceller.
Hos planter er det en generasjonsveksling mellom en diploid sporofyttfase (2n), og en haploid gametofyttfase (n). Sporofyttfasen utvikler seg fra en zygote og produserer sporemorceller som via meiose gir sporer. Hos primitive planter er gametofytten stor, men blir mindre og mindre etter hvert som planteriket utvikler seg, for til slutt å bli mikroskopisk. Hver organisme har en standard karyotype dvs. den artskarakteristiske samlingen av kromosomer i en cellekjerne.
Polyploidi er vanlig hos planter, og refererer seg til antall kopier av kromosomet. Det skjer en genomduplisering ved polyploidi. Polyploidi kan være fordel for plantene siden det reduserer eller eliminerer infertilitet hos hybrider. Polyploidi gir økt volum av cellekjernen, økt antall kromosomatskillelser i meiosen og gjør at celledelingen tar lenger tid. Imidlertid vil økt antall kromosomer gi større muligheter for homolog rekombinasjon med derav følgende genetisk variasjon.
En tetraploid plante har 4 kopier av hvert kromosom i de somatiske cellene. Potet har 48 kromosomer og er en tetraploid plante (2n=4x=48, x=12). I Peru finnes ville slektninger til potet som er haploide med 12 kromosomer. Potet er autotetraploid fordi den dobler kromosomtallet av egne kromosomer, altså en autopolyploid. Polyploidi kan også oppstå ved seksuell krysning av to arter. Allopolyploidi finnes f.eks. hos tobakk (Nicotiana tabacum) som fremkommer ved krysning av N. tomentosiformis med 12 kromosomer og N. sylvestris med 12. Dette gir en hybrid med 24 kromosomer. Brødhvete er heksaploid (2n=6x=42, x=7). Bregner kan ha opptil 1000 kromosomer. Mange planter har imidlertid mange, små og like kromosomer som det er vanskelig å identifisere.
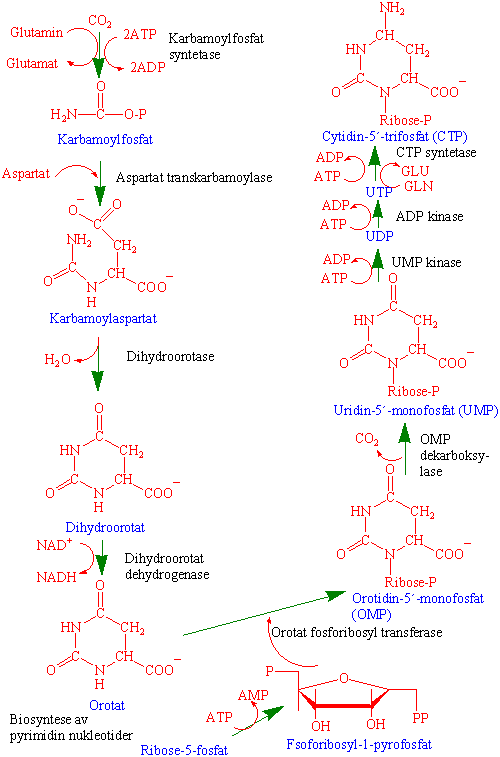
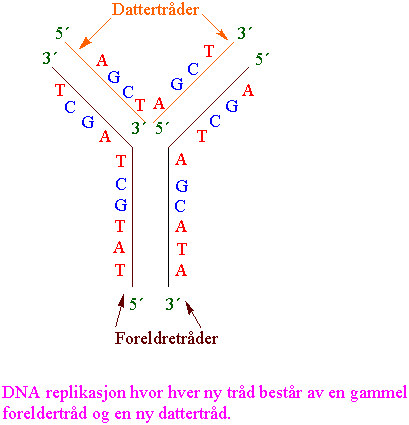
Replikasjon - fra DNA til DNA
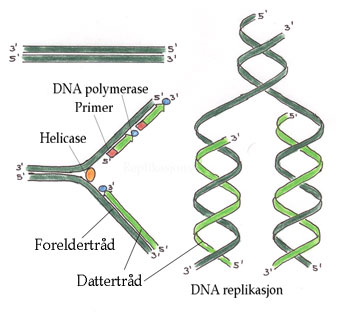
Mellom celledelingene lages et duplikat av hvert kromosom. De to datterkromosomene er i nær kontakt etter kromosomreplikasjonen er fullført, og kromosomet består i dette tilfellet av to kromatider. Kromatidene blir ikke kalt kromosomer før de er separert under mitosen. Mitose fordeler nøyaktig de to kromatidene til de to dattercellene. Under mitosen vil hver datterkjerne motta et komplett sett med kromosomer slik at mitosen endrer ikke kromosomtallet.
Dette i kontrast til meiose hvor reduksjonsdelingen leder til reproduktive celler (kjønnsceller) med halvert kromosomtall. Kromosomet replikeres semikonservativt dvs. hver tråd brukes som oppskrift (templat) for syntese av en kopi.
Første trinn i DNA replikasjonen er opptvinning av DNA-heliksen. Det lages en replikasjonsgaffel hvor enzymene helikase og topoisomerase tvinner opp DNA. Nye kopier av hver atskilt tråd lages av DNA polymerase. Hos eukaryotene er det fire forskjellige DNA polymeraser (a, b, d, e). I mitokondriene er det bare en g-DNA polymerase. DNA polymerase lager en ny tråd fra et lager med deoksyribonukleosidtrifosfater (dNTP).
{kind=link}
Replikasjon av DNA skjer bare i 5´til 3´ retning. Den ene tråden kan derved lages kontinuerlig katalysert av DNA polymerase I. Den andre tråden må lages baklengs i småstykker (Okazaki fragmenter) som limes sammen av enzymet DNA ligase.
Et primase-polymerasekompleks starter DNA syntese av ledetråden med en RNA-primer. Etter start overtas DNA-kopieringen av et nytt polymerasekompleks, og primase-polymerase deltar i dannelser av Okazaki-fragmenter på forsinkelsetråden. Hva som skjer med nukleosomene under replikasjonen er noe usikkert.
Tandemrepetert DNA
Tandemrepetert DNA er korte sekvenser med repetert DNA. Mange korte nukleotidsekvenser kan være repetert fra noen få til millioner av ganger i genomet. Repeterte sekvenser endrer og utvikler seg raskt i genomet i forhold til de kodende sekvenser.
Repetert DNA kan deles i grupper avhengig av lengden:
1) Satelitt DNA.
2) Minisatelitter. Minisatelitter (VNTR loci - variabelt antall med tandem repeterte sekvenser) finnes hos eukaryotene. Hver repetert sekvens inneholder en kort G-rik kjerne varierende fra 11-60 basepar. Minisatelittene er svært variable. Nye varianter oppstår med en frekvens opptil 2% per gamet per generasjon, sammenlignet med gener som har mutasjonsrate 10-5 per generasjon. Det gjør disse viktige som molekylære markører. Disse kan brukes til å lage en DNA profil.
3) Mikrosatelitter. Mikrosatelitter har korte tandemrepeterte polymorfier.
En type repeterte sekvenser koder for produkter som cellen trenger store mengder av f.eks. ribosomalt RNA, organiseres i tandemklynger.
Kloroplastgenomet
Organellene kloroplaster og mitokondrier er etterlevninger etter organismer som inngikk symbiose med forløperene til dagens eukaryoter. Disse organellene kan ikke lenger leve utenfor den eukaryote celle, men innehar fremdeles reminisenser fra deres prokaryote opphav. Primitive eukaryoter som mangler kloroplaster og mitokondrier kan bare leve i et anaerobt miljø. Genomet til disse organellene er små sammenlignet med kjernegenomet og varierer fra 70-400 kbp i kloroplasten. Kloroplast DNA fra alger er ofte større enn de man finner hos planter og disse kan inneholde gener som plantene har lokalisert til cellekjernen. Kloroplasten hos Euglena har tandemrepeterte rRNA gener, noe som ikke finnes hos planter. Genene i kloroplastene ligger i operoner som hos bakterier. En DNA-tråd lager polycistronisk mRNA med en promoter og flere gener med forskjellige funksjon styrt av en promoter. Den motsatte DNA-tråden lager monocistronisk mRNA.
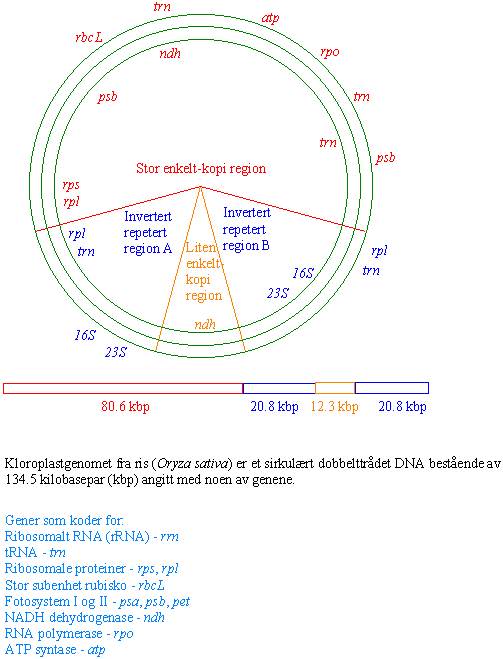
Kloroplast DNA er et sirkulært kromosom bestående av en dobbelhelix uten histoner. Kloroplastgenomet er inndelt i 4 områder: Et stort område med enkeltkopigener og et lite område med enkeltkopigener, og disse to er atskilt av to områder med inverterte repeterte sekvenser. Mye av størrelsesvariasjonen i det sirkulære kloroplastgenomet skyldes to inverterte repeterte sekvenser som atskilller stor og liten enkeltkopiregion. Hos noen kan de inverterte repeterte sekvensene mangle helt, eller være snudd i forhold til hverandre. På utsiden av det inverterte repeterte DNA forekommer hvert gen bare en gang i genomet. Både kloroplaster, kromoplaster, og amyloplaster har alle kopier av det samme kloroplast DNA, dvs. de er genetisk like. Det betyr at gener for fotosyntese også finnes i plastider i rota. Antall kopier av kloroplastgenomet varierer med type celle og grad av differensiering. I mesofyllcellene inneholder kloroplastene 20-40 kopier av kloropastgenomet. Mange kopier av det sirkulære kpDNA i en celle kan dekke et stort behov for proteinsyntese. Imidlertid finnes flere av fotosyntesegenene i cellekjernen bl.a. genet rbcS som koder for den lille subenheten til rubisko. Amyloplasten i potetknoller og kromoplastene i tomater inneholder bare noen få kopier av kloroplastgenomet. Kloroplast-DNA fra levermosen Marchantia og tobakk er omtrent identiske, selv om for til disse artene skilte lag for 100 millioner år siden. Dette viser at kloroplastgenomet endrer seg sakte. Mange av kloroplastproteinene lages i cytoplasma og transporteres inn i kloroplastene. Kloroplast-DNA (kpDNA) inneholder ca. 90 gener som koder for rRNA tRNA, subenheter av ATP syntase, den store subenheten til rubisko, RNA polymerase, fotosystem I og II proteiner og elektrontransportproteiner. Genene transkriberes med klokken i sirkelen. Mange gener er overført fra kloroplasten til kjernen. Det skjer genoverføring mellom organeller og cellekjernen. Man har funnet sekvenser som koder for den store subenheten til rubisko i mitokondriegenomet hos noen arter, men kloroplastgenomet er relativt beskyttet mot å ta imot DNA.
Mitokondriegenomet
Sammenlignet med vertebratene med et mitokondriegenom på ca. 16 kbp, og sopp fra 18-80 kbp, er genomet hos planter stort og meget varierende i størrelse fra 200-2.600 kbp. Størrelsen kan variere selv innen en plantefamilie, men mtDNA koder for relativt få gener (ca. 30) sammenlignet med størrelsen. Variasjonen skyldes størrelsen på ikke-kodende områder mellom genene. Mitokondrie DNA inneholder sekvenser som finnes i kpDNA, men ikke vice versa. Sirkulært mitokondrie-DNA (mtDNA) følger den universelle genetiske koden, og flere promotere som samvirker kan gi forskjellig styrke av et genuttrykk. Mitkondre DNA hos Chlamydomonas er lineært. Det finnes også mindre sirkulære mtDNA i celle laget fra større mtDNA. Disse mindre subgenomiske mtDNA lages ved homolog rekombinasjon fra direkte og inverterte repeterte sekvenser. Både kloroplaster og mitokondrier nedarves via moren, fordi disse organellene elimineres fra cytoplasma under celledelingen som gir spermceller. Mer enn 90 % av artene gjør dette. Også noen typer variegeringer nedarves på samme måte. Mais og flere andre planter har cytoplasmatisk hannsterilitet hvor pollenknappene aborteres eller pollen ikke utvikles. Denne egenskapen nedarves maternalt via mitokondriene.
DNA innhold og C-verdi paradoks
Mange av sekvensene i DNA er repetert en rekke ganger, slik at mengde DNA ikke nødvendigvis gjenspeiler hvor kompleks en organisme er. En organisme kan ha et innhold av DNA som er mye større enn det som trengs for å lage alle proteinene organismen er laget av. Nukleinsyreinnholdet i virus er i størrelsesorden 10.000-100.000 basepar (bp) og for bakterier ca. en million basepar. Mennesket med 3.2 milliarder basepar har altså mer enn 1.000 ganger mer DNA enn bakteriene. Alger kan ha 100 ganger mer DNA enn høyere planter. Det er dette som kalles c-verdi paradoks, og som skyldes polyploidi og repeterte sekvenser. Mengden DNA i haploid generasjon kalles genomstørrelsen eller C-verdi. C-verdien angis i pikogram (pg), som antall basepar eller i dalton. 1 pg er 10-12 g= ca. lik 109basepar = ca. 6.4×1011 dalton. 1 pg DNA er ca. 31 cm langt og inneholder ca. 109 basepar. I eukaryoter angir C-verdien som mengde DNA per genom som 1C for haploide kjønnsceller og 2C for diploide kjerner. C- verdien er større for eukaryoter enn prokaryoter. Arabidopsis som har det minste kjente plantegenom har bare ca. 10% av DNA som repeterte sekvenser i ikke-kodende introns, mens andre planter kan ha over 80 % av DNA i cellene som repeterte ikke-kodende sekvenser. Det er mulig at repeterte sekvenser i DNA har en strukturfunksjon i kromosomene.
Vårskrinneblom (Arabidopsis thaliana) har ca. 0.07 pg DNA per haploid celle, sammenlignet med hestekastanje (Aesculus hippocastanum) som er et stort tre og har ca. 0.3 pg DNA. Mais som har 100 x så stort genom inneholder ikke 100 ganger så mange gener. På en todimensjonal akrylamidgel kan opptil 2000 proteiner identifiseres ved å gi flekker etter farging. Det er imidlertid få proteiner som forekommer i store mengder i en celle. Planten har 8-12 forskjellige organer som vil gi ca. 8000-12000 organspesifikke gener, ialt ca. 28000- 32000 gener. Omtrent halvparten av plantene har polyploidi. Familier av repeterte sekvenser har utviklet seg fra eldre former. Mengden repetert DNA står for en stor del av variasjonen av genomstørrelse. Det finnes fra 100-40 000 forskjellige repeterte DNA-familier. Noen er tandem repeterte og andre er spredde. Repetert DNA har sannsynligvis betydning for evolusjon og artsdannelse.
Opptil 10% av DNA kan være transposoner og det er mange kopier per celle. Et transposon kan bestå av fra 100-1000 basepar og flyttes rundt i genomet katalysert av en transposase.
Kjernegenomet og kromosomer
Organismene har vanligvis to sett (utgaver) av hvert gen som nedarves via Mendelsk forhold. En celle med to sett av kromosomer kalles diploid. Gametene sies å være haploid og den spesielle celledelingen som halverer kromosomtallet kalles meiose. Befruktning av egget med sperm resulterer i en diploid zygote. Plasseringen av et gen på kromosomet kalles et locus, og et individ har derved 2 gener ved hvert locus. Ett gen kommer fra faren og ett fra moren. De to genene i hvert locus kalles en genotyp. Hvis de er like kalles genotypen homozygot, og er de forskjellige kalles genotypen heterozygot. Forskjellige utgaver av et gen i et locus kalles alleler (genutgaver). Med 2 alleler i et locus er det f.eks. 3 mulige genotyper AA, Aa og aa. Genene påvirker egenskapene til et individ (fenotype). F.eks. kalles allelet A, angitt med stor bokstav, for dominant og a med liten bokstav kalles recessivt. Et allel er dominant hvis heterozygoten har samme fenotyp som homozygoten for vedkommende allel. Mendelforholdet uttrykker forholdet mellom de forskjellige genotypene i avkommet fra foreldre med en spesiell kombinasjon av genotyper. Den enkleste kryssingen er mellom AA hunn og en AA hann hvis vi bare ser på ett gen. Da blir det 100 % avkom med genotypen AA. Krysser vi derimot AA og Aa vil vi få genotypene 25 % AA, 50 % Aa og 25 % aa. Atskillelse av 2 gener i et locus kalles segregasjon (sortering). En krysning mellom hann AB/Ab og hunn AB/ab vil
gi genotypene 1/8 AB/AB, 1/4 AB/Ab; 1/8 Ab/Ab; 1/8 AB/aB; 1/4 AB/ab; 1/8 Ab/ab. Segregasjon av ulenkete genotyper kalles uavhengig sortering (segregasjon). Gener på et kromosomer kan være lenket sammen. Hvis loci er lenket på samme kromosom blir de ikke sortert uavhengig av hverandre. I meiosen kan det skje rekombinasjon, de to trådene kan heftes sammen og det skjer bytte av basesekvenser. Rekombinasjon er en tilfeldig prosess med sannsynlighet r. Sannsynligheten for ikke rekombinasjon er 1-r. Forholdet mellom genotyper i avkommet for to lenkete gener avhenger av rekombinasjonen mellom dem. Store genomer har ofte metylert cytosin laget ved metylering. Mange gener er beskyttet mot metylering. I planter kan opptil en tredjedel av cytosin kan være metylert. Metylering kan påvirke transkripsjonen.
Kromosomet består av en lang DNA heliks tvunnet rundt nukleosomer (nukleoproteinkjeder). Enden av kromosomet har korte repeterte sekvenser kalt telomere som beskytter enden av kromosomet under replikasjonen. I metafase er kromosomene mest kondensert og lettest å observere cytologisk.
Gjennomføring av en cellesyklus med mitose hos planter kan ta fra 10-24 timer, mens meiosen kan vare fra 24 til 280 timer, kanskje lenger. Hvordan kromosomene oppfører seg under mitosen kan studeres ved å bruke cellevev som deler seg aktivt f.eks. rotspisser. Kromosomene farges med en basisk farge, Feulgen-reagens, som reagerer med aldehydgrupper i DNA etter en mild sur hydrolyse. Et synaptonemisk kompleks dannes mellom homologe kromosomer og kan sees i elektronmikroskop etter en spesiell fargeprosedyre. Det er sannynlig at det er her rekombinasjoner foregår. Chiasmata er stedet hvor det skjer fysisk brudd og reforbindelse (rekombinasjon) mellom søsterkromatider. Chiasmata kan sees i lysmikroskop Polyploide har mer enn to komplette sett med kromosomer. De kan enten være autopolyploide, hvor alle kromosomene kommer fra samme art, eller allopolyploide, hvor det ekstra kromosomsettet, som enten kan være homologt eller ikke, kommer fra en annen art. P.g.a. de ekstra kromosomsettene kan det oppstå problemer under atskillelsen av kromosomene under reduksjonsdelingen slik at kjønnscellene ikke får med seg fullt sett av kromosomene. F.eks. for en trivalent (2n=3x) så vil bare to kromosomer kunne danne par. Dette betyr at resultatet kan bli en univalent og en bivalent, eller en trivalent. Triploider danner alle typer konfigurasjoner i første metafase, og ubalanserte gameter gir sjeldent fertilt avkom. Parvis polyploidi som f.eks. tetraploider er mer fertile enn ikke- parvise.
Inversjoner skjer når det er to brudd på et kromosom og segmentet mellom de to brudd reorienteres i motsatt retning og kobles sammen på nytt. Ved delesjoner fjernes et segment av kromosomet. Translokasjoner er bytte av gensegmenter mellom ikke-homologe kromosomer. Genkart sier noe om plasseringen av genene på kromosomene ut fra meiotiske rekombinasjonsdata. Cytologiske genkart sier noe om den fysiske plasseringen av et gen på et kromosom. Genkart, bindingsanalyse, baseres seg på Mendels andre lov om uavhengig sortering av alleler under dannelse av kjønnscellene. Det var T.H. Morgan som i 1911 foreslo at styrken av binding mellom to gener avhenger av avstanden mellom dem på kromosomet. Jo større avstand desto større sannsynlighet for rekombinasjon mellom genene. Slike data skaffer en seg ved å krysse organismer som gir kvantifiserbare forskjeller i avkommet. Aneuploide har ikke normalt diploid (eller euploid) kromosomtall. Den vanligste typen er monosomi (2n= 2x-1) som mangler et kromosom, og trisomi (2n=2x+1) som har et ekstra kromosom. Trisomi kan man få ved å krysse en diploid med en tetraploid. Man antar at ca. 70 % av alle planter er hermafroditter. For å motvirke selvpollinering er det forskjellige mekanismer som at kjønnsorganene utvikler seg på forskjellig tid (asynkrone hermafroditter). Selvpollinering kan hindres ved cytoplasmatisk hannsterilitet og selvinkompatibilitet. Andre mekanismer som sikrer krysspollinering er forskjellige hann- og hunnblomster på samme plante (monoik) eller på forskjellige planter (dioik). Kjønnskromosomer forekommer sjelden hos planter. Av planter hvor man kjenner til kjønnskromosomer er humle (Humulus lupulus, rød og hvit jonsokkblom (Silene dioica og S. latifolia alle med XX-XY-systemer, og engsyre (Rumex acetosa) med XX-XY1Y2. Hos hvit jonsokkblom er hunnplantene 2n=22A + XX og hannplantene har 2n=22A + XY (A- autosom).
Kosuppresjon
Fenomenet kosuppresjon ble oppdaget da man forsøkte å få kraftigere farge på blomstene av petunia (Petunia hybrida) ved å sette inn et ekstra gen for chalkon syntase. I motsetning til det som var forventet ble resultatet hvite områder på kronbladene. To eller flere homologe gener ble stille og satt ut av drift. Det viste seg at genene ble transkribert, men mRNA ble raskt nedbrutt slik at det ikke ble laget noe protein. Transgen tomat med redusert innehold av polygalakturonase ble laget med denne teknikken ved å sette inn et gen for polygalakturonase. Dette kan var en mekanisme for å beskytte seg mot og bekjempe virus. Kosuppresjon kan komme etter en periode med stress.