- Historien om abscisinsyre
- Generelt om abscisinsyre
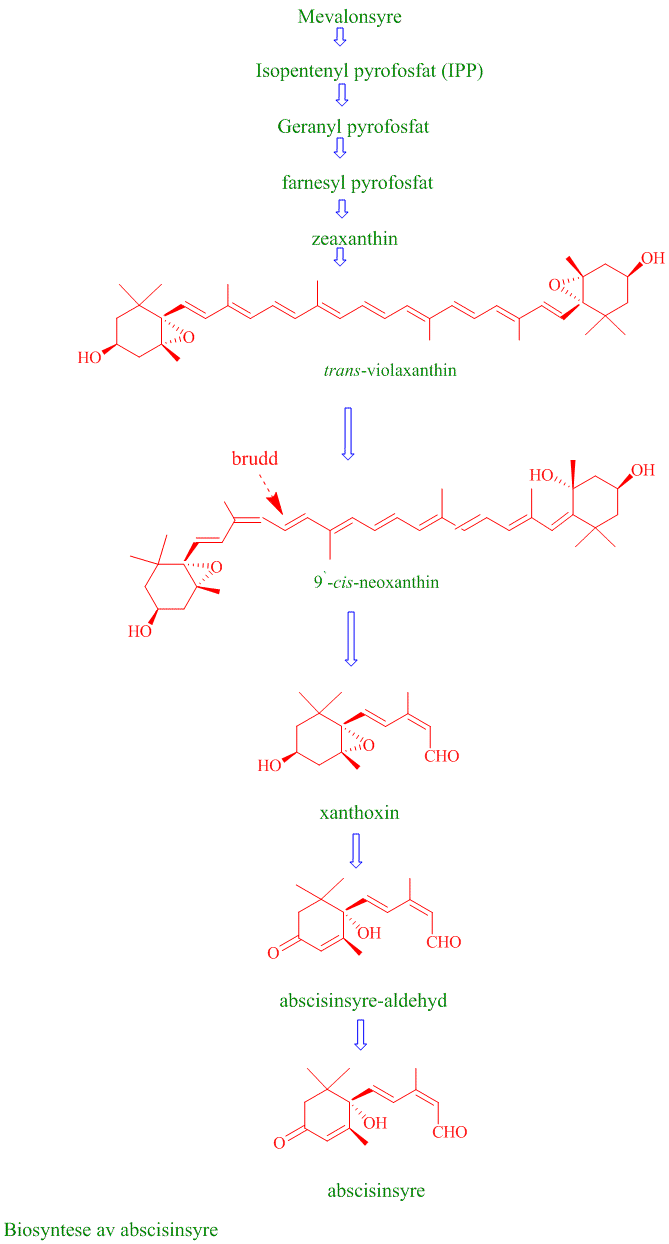
- Biosyntese av abscisinsyre
- ABA og tørkestress
- Abscisinsyre og vivipari
Historien om abscisinsyre
I lang tid mente man at knopphvile og frøhvile skyldtes hemmende stoffer. Liu og Carns i 1961 isolerte et stoff fra modne bomullsfrukter som stimulerte felling (avkastning, abscission) av bomullsbladstilker. De kalte stoffet abscisin I. Seinere ble stoffet isolert som en 15- karbonforbindelse kalt absicisin II. Omtrent samtidig isolerte Warings gruppe i England en veksthemmende substans fra bjerkeblad eksponert for korte dager. Stoffet hemmet vekst av den apikale knoppen på frøplanter av lønn (Acer). De antok at det var et dvaleinduserende stoff og kalte det dormin. Etterhvert ble man enige om å kalle stoffet abscisinsyre. Abscisin II var det samme som dormin og absiscinsyre (ABA= abscisic acid). Dormin hadde vært et mer velegnet navn med tanke på ABA og frøhvile, mens ABAs rolle ifm. absisjon er mer indirekte, hvor flere andre hormoner deltar.
Generelt om abscisinsyre
Abscisinsyre og tilhørende signalveier er viktige i evolusjonen av planter og moser, med tilpasning til et liv på land med tørkestress. Abscisinsyre (ABA) har lite med bladavkasting å gjøre og har derved et noe misvisende navn. ABA gir avkastning av bomullsfrukt ved å stimulere etylensyntese. ABA ble opprinnelig funnet i blad av bjerk og lønn og i bomullsfrø. ABA finnes i hvilende knopper og frø, og deltar i lukkingen av spalteåpningene ved vannmangel. Konsentrasjonen av ABA øker i hvilende knopper selv om konsentrasjonen av ABA ikke er korrelert med hvile.
Mange andre veksthemmende stoffer er kjent fra planteriket. Mange av disse finnes i frø. Noen beskytter frø som blir liggende flere år i jorda før de får anledning til å spire. Andre hindrer frøene å spire med en gang de er modne. ABA lages under forskjellige typer stress forårsaket av kulde, salt (saltstress), og vannmangel (tørkestress). Blad med forskjellig form over og under vann kan skyldes ABA. ABA hemmer cellestrekning i skuddvev, induserer lukking av stomata, fremmer avkasting av blad og frukt og øker eller induserer hvile i frø og knopper. ABA hemmer dessuten DNA syntese, aktiverer ribonukleaser, og øker membranpermeabilitet. ABA hemmer gibberellinindusert syntese av enzymet a-amylase.
ABA fraktes lett rundt i planten i silvev (floem) og vedvev (xylem). ABA kan fraktes fra rota til skuddet.
Biosyntese av abscisinsyre
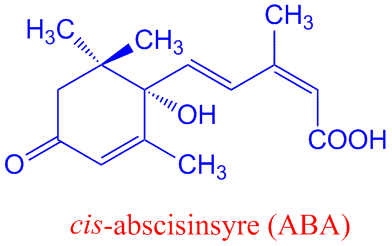
Abscisinsyre er et sesquiterpenoid, bestående av 15 karbonatomer i en alifatisk ring med en dobbelbinding, 3 metylgrupper og en umettet sidekjede. Naturlig forekommende abscisinsyre (ABA) er en cis-isomer ved karbonatom 2 i sidekjeden. ABA har også et assymetrisk karbon i 1´- posisjon på ringen som gir mulighet for S(+) og R(-) enantiomere. Det er S-enantiomeren som er naturlig forekommende. ABA lages fra xanthofyll i kloroplaster og plastider via mevalonsyresynteseveien. ABA finnes i hele planteriket, moser (untatt levermoser), grønnalger, og sopp, men har ikke blitt funnet i bakterier. Det er mulig at lunularsyre har funksjonen til ABA i levermoser. ABA lages via mevalonatveien enten fra C15 farnesylpyrofosfat eller fra spalting av C40 karotenoid (vioalaxanthin) i kloroplastene. Violaxanthin blir laget fra zeaxanthin katalysert av zeaxanthin epoksidase. Violaxanthin omdannes til 9´-cis-neoxanthin, katalysert av enzymet 9-cis-epoksykarotenoid dioksygenase (NCED), og videre til 15-karbonforbindelsen xanthoxin. Til slutt dannes ABA via oksidasjon og intermediatet ABA-aldehyd katalysert av abscisinsyre-aldehyd oksidase, med molybden som en kofaktor. Enzymet NCED blir kodet av en multigenfamilie, med vevsspesifikke uttrykk i røtter, lukkeceller, eller vaskulært parenkym, og enzymet blir indusert av tørkestress.
Konjugering og inaktivering av ABA
ABA kan oksideres til 6-hydroksymetyl-ABA som omdannes videre til phaseinsyre og dihydrophaseinsyre. ABA kan glukosyleres og konjugeres til monosakkarider som f.eks. ABA-\(\beta\)-D-glukosylester som akkumuleres i vakuolen, mens ABA finnes i cytosol. ABA kan bli inaktivert via hydroksylering av karbonatom 7, 8 eller 9, og videre inaktivering via phaseinsyre og dihydrophaseinsyre. Inaktiveringen skjer etter at tørkestresset er fjernet, og planten får god vanntilgang
{kind=link}
ABA og tørkestress
ABA (abscisinsyre) deltar i åpning og lukking av stomata, fungerer som tørkestresssignal og induserer forsvar mot saltstress og kuldestress. Tørkestress kan gi opptil 50 x økning i konsentrasjonen av ABA. ABA er en svak syre, med pH-likevekt mellom ladet ABA- og og uladet ABAH, hvor sistnevnte lett kan passere membraner. Ladet ABA- blir sannsynligvis fraktet over membranen via ABC-transportører. Ved tørkestress stiger pH i xylemet fra ca. 6.3 til 7.2 og dette gir en dissosiert form av ABA, ABA-, som lettere kommer via apoplasten fram til lukkecellene. Tørketilstand med lavere tilgjengelighet av vann blir først registrert av røttene hvor det skjer økt syntese av ABA. Økt ABA-konsentrasjon i røttene gir økt rotvekst. Noe ABA blir fraktet til skuddsystemet via xylemet. Konsentrasjonen av ABA i bladet øker kraftig ved tørke, noe som også skyldes nysyntese i bladet.
ABA deltar under normal embryogenese og konsentrasjonen av ABA øker kraftig i en liten periode under frøutvikling. ABA deltar også i syntese og akkumulering av frølagringsproteiner og lipider.
Første fasen av embryogenesen er celledeling. I den andre fasen stopper celledelingen og det skjer en uttørking av embryo som går inn i en hvilefase (frøhvile) hvor vannpotensialet kan bli -50 - -300 MPa. Frøhvilen kan skyldes uferdig embryo, frøbladene, frøskallet, endosperm, eller perikarp. Vannopptaket kan hindres på grunn av voks eller frøskallet kan være hardt og hindre frøspiring mekanisk. Frøskallet kan dessuten hindre gassutveksling. Det kan være spiringshemmere i frøskallet. Frøskall og perikarp kan også lage spiringshemmende stoffer. Når frøet går ut av frøhvile synker konsentrasjonen av ABA, mens konsentrasjonen av gibberellin øker. Faktorer som bidrar til avslutning av frøhvilen er ettermodning ved tørking eller kjøling i fuktig miljø ved 0-10 oC (stratifisering). Lys kan påvirke rotas evne til å trenge gjennom frøskallet.
Det er zygoten som bestemmer mengden ABA i embryo og endosperm, mens det er morplanten som bestemmer mengden ABA i frøskallet. ABA er med å gi frøet tørketoleranse. Det lages LEA- proteiner (late embryogenesis abundent protein) som er vannløselige og inneholder mye av aminosyrene glycin og lysin. LEA-proteiner ligner på RAB-proteiner ("responsive to ABA") og DHN-proteiner (dehydrin) i Arabidopsis.
ABA øker ionefluksen og den hydrauliske konduktivitet i røtter. Det vil si at effekten av ABA skjer ikke bare via spalteåpningene.
ABA øker veksten av røtter og hemmer veksten av skuddet ved lave vannpotensialer. ABA kan dessuten fremme bladaldring uavhengig av etylen.
ABA regulerer genekspresjon av flere gener og det finnes abi-mutanter av Arabidopsis ("ABA insensitive") hvor produkter fra abi-gener ikke bare påvirker frøutvikling og frøhvile, men også vegetativ vekst. Genaktiveringen skjer via DNA-bindende proteiner. ABA forårsaker at Ca2+ i cytosol øker kraftig. pH øker også, og dette gir en depolarisering av membranpotensialet med rask innstrømning av kationer og endret metabolisme av fosfoinositider (inositol, 1,4,5-trifosfat, IP3).
Lite vann og lite nitrogen leder til økt syntese og eksport av abscisinsyre fra røttene til skuddet. I skuddet kan abscisinsyre påvirke cellemembraner og gi lukking av spalteåpningene. Tilførselen av abscisinsyre stopper skuddveksten, men ikke rotveksten. Nitrogenmangel gir økt rotvekst og dette kan delvis forklares som en effekt av abscisinsyre. Redusert rotvekst for planter med en liten potte resulterer i redusert skuddvekst, ikke bare på grunn av mangel på vann og næringssalter, men fordi at det sendes et stressignal til skuddet. Ved nitrogenmangel synker innholdet av gibberellin, mens mengden abscisinsyre øker. Unge ekspanderende blad trekker sterkt på fotosynteseproduktene. Kunstig tilføring av abscisinsyre reduserer bladekspansjonen og øker eksporten av fotosynteseprodukter fra bladet. Abscisinsyre virker således ved å skifte fra import til eksport av fotosynteseprodukter fra et blad.
Hos tomat finnes det flc-mutanter (flacca) som tørker ut og disse kan ikke åpne spalteåpningene.
Selv om plantehormonet etylen er viktig ved modning av frukt, så deltar også ABA ved modning av både klimakterisk og ikke-klimakterisk frukt
Abscisinsyre og vivipari
Vivipari finnes hos mange frø som mangler abscisinsyre. Mutanter som har blokkert karotenoidbiosyntesen er ofte vivipare. Vivipari er naturlig hos mangrove (Rhizophora). Vivipari er også vanlig hos fjellplanter. Mais vp1 er en viviparmutant (viviparous) hvor embryo ikke går i hvile. vp1 har også pleiotrope effekter på anthocyaninsyntese under frøutviklingen hvor anthocyaninsyntese i aleuronlagceller blokkeres. Genproduktet VP1 er sannsynligvis en transkripsjonsfaktor som påvirker flere gener.