{kind=link}

Xanthofyllsyklus (VAZ-syklus) fjerner overskuddsenergi som varme
Karotenoider som ligger i tylakoidmembranene kan både virke som lyshøstende pigment (absorberer lysenergi fra 450-500 nanometer (nm) som overføres til klorofyll), men har også en funksjon i fotobeskyttelse av fotosynteseapparatet. Karotenoidene virker som en sikkerhetsventil som fjerner overskudd av eksitasjonsenergi som kommer inn i fotosyntesen ved høy lysfluks. Karotenoidene virker i denne fotobeskyttelsen ved å fjerne eksitasjonsenergi fra klorofyll som varme. Ved ikke-fotokjemisk quenching føres lysenergi vekk fra elektrontransport over til varmeproduksjon.
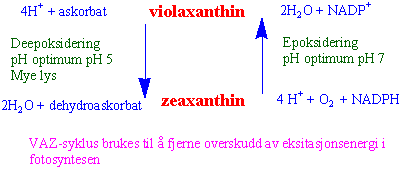
Xanthofyllsyklus er en syklus som gir en lysavhengig og reversibel minskning av violaxanthin i grønne blad. Violaxanthin omdannes til zeaxanthin via intermediatet antheraxanthin. VAZ-syklus fjerner overskudd av eksitasjonsenergi som varme og beskytter planten mot fotooksidative skader under lysstress. Enzymet som katalyserer overføringen av violaxanthin til zeaxanthin er en del av tylakoidmembranen inn mot lumen og har et pH optimum ved pH 5.2. Under elektrontransporten i fotosyntesen pumpes protoner (H+) inn i lumen på tylakoidmembranne. Hvis syntesen av ATP går saktere så øker konsentrasjonen av H+ inne i lumen og mengden zeaxanthin øker. Enzymet de-epoksidase overfører violaxanthin til zeaxanthin med askorbat som kofaktor. Hvis derimot ATP syntesen er høy blir det mindre pH gradient og reaksjonen skjer fra zeaxanthin til violaxanthin katalysert av en epoksidase som bruker NADPH.
Mye lys, lav CO2, lav pH i lumen gir følgende reaksjon for H+ og askorbat (vitamin C) deltar:
violaxanthin → antheraxanthin→ zeaxanthin
Lite lys, høy CO2 , høy lumen pH hvor NADPH, oksygen og H+ deltar:
zeaxanthin → antheraxanthin → violaxanthin
Det er indikasjoner på at det kan være en sammenheng mellom konsentrasjonen av zeaxanthin og åpning og lukking av spalteåpninger. Kloroplasten i mørkespirte koleoptiler inneholder violaxanthin og antheraxanthin, mens i lys dannes zeaxanthin. Dette kan muligens skyldes at kloroplastene i koleoptilen har en spesiell funksjon.
Blad i fullt sollys har høyere konsentrasjon av VAZ metabolitter enn i skyggeblad, og i mye lys utgjør zeaxanthin og anteraxanthin 60% av poolen med VAZ. Det finnes imidlertid også VAZ i skyggeblad, og dette virker ved lysglimt som kommer ned til eller skyggefull skogbunn. VAZ-syklus er viktige i vintergrønne bartrær ved lav temperatur og mye sollys. Utskillelsen av isopren er større i sol enn i skygge, men det skjer nysyntese av isopren ved høy temperatur.
Tilbake til hovedside