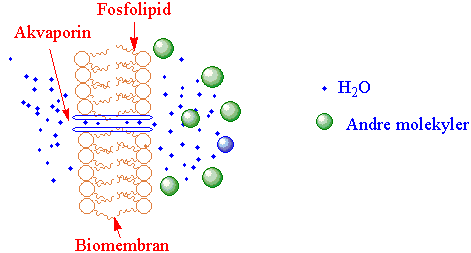
Vanndiffusjonen gjennom membraner blir styrt av membrankanalproteiner (akvaporiner).
Peter Agre fikk nobelprisen i kjemi 2003 for oppdagelsen av akvaporiner.
Aminosyrer med positiv ladning i kanalen gjør at hydroniumioner (H3O+) ikke passerer gjennom akvaporiner. Protongradienter har viktige funksjoner, og disse blir ikke forstyrret av akvaporinene.
Vann som blir tatt opp av røttene blir forflyttet og fordelt i planten avhengig av en vannpotensialgradient. Akvaporiner er en heterogen gruppe membraner som kanaler gjennom cellemembraner og regulerer transporten av vann over membranene og påvirker hydraulisk konduktanse. Akvaporiner kan ut fra sekvensdata deles i familier fordelt på plasmamembranen (cellemembranen), vakuolemembranen (tonoplast), endoplasmatisk retikulum og peribakteroidmembranen hos nitrogenfikserende erteplanter. Akvaporinene finnes i stort antall og er med å opprettholde likevekt (homeostase) i vanninnhold i planten underskiftende utviklingsstadier og varierende ytre miljøbetingelser, inkludert osmoregulering. Det er ikke bare vann som kan passere membranen via akvaporiner, men også uladete molekyler som karbondioksid (CO2), urea, ammoniakk (NH3), borsyre, kiselsyre, glycerol og hydrogenperoksid (H2O2). Tettheten av vannkanaler i membranen kan variere med de ytre forhold, og vannkanaler kan bli fjernet fra membranen via endocytose. Åpningen av vannkanalen blir regulert av divalente ioner som kalsium (Ca2+), pH i cytoplasma, abscisinsyre, metylering av aminosyrer, fosforylering via protein kinaser og defosforylering katalysert av fosfataser, som kan påvirke spesifikke aminosyrer i kanalen. Vannkanalene blir nedregulert ved tørke og minralnæringsstress. For eksempel vil mangel på oksygen (anoksis) gi økt surhet i cytoplasma og redusert vanntransport. Syntesen av vannkanalproteiner følger cirkadiske rytmer og har posttranslasjonsregulering. I parenkymceller tilknyttet ledningsvevet er det mange vannkanaler. Størrelsen av proteinet med seks transmembrandomener er ca. 30 kD. Både den C- og N-terminale delen av proteinet befinner seg i cytosol. Løkke I, III og V av det foldete proteinet er i apoplasten, og løkke II og IV er seg i cytosol.
Akvaporiner hos dyr
Akvaporiner er proteinkanaler gjennom membraner, og noen av dem kan i tillegg til vann frakte glycerol. Hos pattedyr er det elleve forskjellige akvaporiner (AQP1-11) e.g.
AQP2: Røde blodceller, nyrer, lunge, hjerne, endotel i blodårer.
AQP3: Øye, tykktarm, spyttkjertler. I tillegg er det akvaporiner i muskler, nyrer, lunger, mage, tynntarm, tykktarm, spyttkjertler, svettekjertler, tårekjertler, bukspyttkjertel, lever, kjønnskjertler, leukocytter. Gjennom et akvaporin kan det fraktes 109 vannmolekyler per sekund avhengig av vannpotensialgradient eller osmotisk gradient.