Sukker lagret som en polymer i glykogen gir få osmotiske effekter sammenlignet med samme mengde fritt sukker. Glykogen kan bestå av opptil 50000 glukosemolekyler. Flere glykogengranuler inngår i rosettformete klynger i cytosol. Glykogen i muskler er et reservoir med energi som raskt kan bli mobilisert, mens glykogenlageret i lever kan gi energi i 1-2 døgn hvis det ikke tilføres ny mat. Deretter må kroppen tære på fettlageret.
Nedbrytning av glykogen
Glykogen blir nedbrutt (glykogenolyse) til glukose-1-fosfat katalysert av enzymene glykogen fosforylase, fosfoglukomutase og glykogenavgreiningsenzym (fjerner greiningspunktene i glykogen).
Glykogen fosforylase spalter α-1,4-glykosidbinginger mellom glukosemolekylene, i en fosforylasereaksjon som er avhengig av vitamin B6 (pyridoksalfosfat).
(α-1,4-glykogen)n + Pi ↔ (α-1,4-glykogen)n-1 + α-D-glukose-1-fosfat
Glykogen fosforylase kapper av enheter med glukose-1-fosfat fra den ikke-reduserende enden av glykogen inntil enzymet treffer på et greiningspunkt med glukose i α-1,6-binding, som brytes av glykogenavgreiningsenzym (en transferase og α-1,6-glukosidase).
Denne reaksjonen er forskjellig fra nedbrytning av glykogen via maten som er en hydrolyse.
For å kunne bli utnyttet i metabolismen i musklene må glukose-1-fosfat omdannes til glukose-6-fosfat katalysert av enzymet fosfoglukomutase:
Glukose-1-fosfat ↔ glukose-6-fosfat
Glukose-6-fosfat kan deretter gå inn i glykolysen og sitronsyresyklus og gi kjemisk energi (ATP) til muskelkontraksjon.
Når sukkerkonsentrasjonen i blodet etter hvert begynner å synke starter nedbrytning av glykogen i lever, hvor leverenzymet glukose-6-fosfatase deltar, hydrolyserer og danner glukose og fritt uorganisk fosfat. Glukose fraktes ut av levercellene via glukosetransportører.
Glukose-6-fosfat blir fraktet via en egen transporter inn i lumen på endoplasmatisk retikulum som ligger i cytosol. ER inneholder glukose-6-fosfatase som spalter glukose-6-fosfat til glukose og uorganisk fosfat. Fosfattrasportøren frakter ut fosfat fra ER, mens glukosetransporter frakter ut glukose. En annen glukosetransporter frakter glukose over plasmamembranen og ut i blodkapillarene. Både oppbygging og nedbrytning av glykogen er under sterk reguleringskontroll blant annet styrt av hormonene glukagon, adrenalin og insulin, samt sekundære budbringer som syklisk AMP (cAMP), AMP, kalsium (Ca2+)-kalmodulin, glukosetransportører, og flere protein kinaser og protein fosfataser. I bukspyttkjertelen (pankreas) roduserer β-cellene insulin, α-cellene glukagon og δ-cellene somatostatin. I tillegg produserer bukspyttkjertelen fordøyelsesvæske. Adrenalin (epinefrin) blir laget i binyrebarken. Glukoseomsetning oppbygning eller nedbrytning av glykogen er også nær koblet til omdanning av glukose til acetyl-CoA og videre til triglycerider (fett i fettvev) med regulering av acetyl-CoA karboksylase . I tillegg er det sentrale reguleringsenzymer i glykose og sitronsyresyklus e.g. glukokinase og pyruvat dehydrogenase. Et samvirke mellom lever og muskler e.g. Corisyklus. Et utrolig vakkert og komplekst biokjemisk regulerrt samspill fintslipt gjennom evolusjon for å kunne holde kontroll med energimetabolismen og energirike molekyler, valg mellom lagring eller omsetning.
Biosyntese av glykogen (glykogenese)
Glukose som inngår i glykogen blir tilført via sukkernukleotidet uridindifosfat-glukose (UDP-glukose). Biosyntesen av glykogen starter med glukose-6-fosfat laget fra glukose katalysert av heksokinase :
D-glukose + ATP → D-glukose-6-fosfat + ADP
Heksokinase finnes som flere isoenzymer, heksokinase I og II i muskler og heksokinase IV i lever.
Fosfoglukomutase katalyserer overgangen mellom glukose-6-fosfat og glukose-1-fosfat:
Glukose-6-fosfat ↔ glukose-1-fosfat
Deretter lages UDP glukose ved å koble sammen glukose og uridintrifosfat (UTP) katalysert av UDP-glukose pyrofosforylase
Glukose-1-fosfat + UTP → UDP-glukose + PPi
Energien som ligger i pyrofosfat PPi blir frigitt katalysert av enzymet pyrofosfatase.
Glykogen syntase er enzymet som overfører glukose fra UDP-glukose til den ikke-reduserende enden av glykogen i alfa-1,4-binding (α-1,4). For å kunne lage alfa-1,6-bindinger (α-1,6) )i greiningspunktene i glykogen brukes enzymet glykogen greiningsenzym, en glykosyltransferase som hekter på ca. 6 glukosemolekyler i greiningspunktet. Glykogen syntase kan ikke starte på bar bakke, men trenger en start (primer) i form av et stort proteinkompleks kalt glykogenin.
Glykogenregulering
Aktivering av glykogen fosforylase
Glykogen fosforylase finnes i to former en aktiv glykogen fosforylase a, og en inaktiv glykogen fosforylase b. Glykogen fosforylase b blir aktivert via fosfosforylering katalysert av glykogen fosforylase b kinase. Signalkaskaden som gir fosforylering av glykogen fosforylase b starter med at glukagon i lever ved lav glukosekonsentrasjon i blodet eller adrenalin (epinefrin) i muskler ved raskt fluktbehov øker omdanning fra ATP til budbringermolekylet syklisk AMP (cAMP) katalysert av adenylyl syklase. Syklisk AMP aktiverer cAMP-avhengig protein kinase A, og det er denne kinasen som aktiverer isoenzymer av glykogen fosforylase b kinase via fosforylering, henholdsvis i lever eller muskler. I tillegg til hormonregulering er det allosterisk regulering av enzymaktivitet. Glykogen fosforylase b kinase blir aktivert av kalsium (Ca2+)- kalmodulin. Kalsium deltar i muskelsammentrekning og når muskler er i aktivt arbeidet er energikilden adenosin trifosfat (ATP), men hvor det også blir laget adenosin monofosfat (AMP). AMP som bindes til glykogen fosforylase gir signal om å øke nedbrytningen av glykogen til glukose-1-fosfat. I glykolysen blir glukose-1-fosfat brukt til arbeid i musklene, mens i leveren blir den brukt til å øke konsentrasjonen av glukose i blodet.Ved resusert muskelbruk blir ikke så mye ATP omsatt, gir økt ATP-konsentrasjon som blokkerer for binding av AMP.
glykogenfosforylase b (inaktiv) + 2ATP+Ca2+ + AMP → glykogenfosforylase a (aktiv) + 2ADP
Inaktivering av glykogen fosforylase
Når musklene går i hvile minsker behovet for nedbrytning av glykogen. Aktiv fosforylert glykogen fosforylase a blir defosforylert, fosfat bundet til hydroksylgruppen i aminosyren serin blir fjernet av en glykogen fosfoprotein fosfatase:
glykogenfosforylase a (aktiv) + 2H2O → glykogenfosforylase b (inaktiv ) + 2 Pi
Glykogen fosfoprotein fosfatase som stopper nedbrytningen av glykogen ved å danne inaktiv glykogen fosforylase b blir også styrt av hormon konsentrasjon, kalsium og AMP, men nå motsatt vei.
Aktivering av glykogen syntase
Glykogen syntase blir aktivert ved at inaktiv og fosforylert glutamin syntase b (fosforylert serin via glykogen syntase kinase 3 (GSK3) blir defosforylert av fosfoprotein fosfatase 1 (PP1) og danner aktiv glykogen syntase a. Imidlertid kan ikke GSK3 fosforylere glykoen syntase b før glutamin syntase a også er fosforylert av kasein kinase II. I tillegg deltar en AMP-aktivert protein kinase. Fosfoprotein fosfatase 1, i tillegg til å aktivere glykogen syntase, blir også aktivert av glukose, glukose-6-fosfat og insulin. Insulin virker også ved å hemme GSK3, og dette skjer ved at insulin aktiverer en protein kinase B som inaktiverer GSK3 via fosforylering. Konsetrasjonen av insulin stiger når det er høy konsentrasjon av glukose i blodet. Glukosetransportøren GLUT2 deltar i å få glukose ut av blodårene ut i cellevevet der det kan bli omdannet til glykogen i muskler eller glykogen og fett i lever.
I bukspyttkjertelen (pankreas) produserer β-cellene i de Langerhanske øyene insulin, α-cellene glukagon og δ-cellene somatostatin. I tillegg produserer bukspyttkjertelen fordøyelsesvæske. Adrenalin (epinefrin) blir laget i binyrebarken, hormonstyring av aktivitet via endokrinologi.
Glukosetransportør GLUT4 i plasmamembran
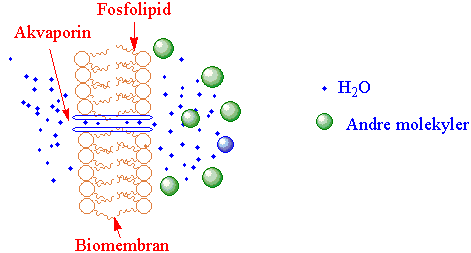
Opprettholde likevekt (homeostase) er sentralt i dyrefysiologi hvor kroppstemperatur, blodtrykk, elektrolyttkonsentrasjon, vanninnhold og glukosekonsentrasjon (ca. 5 millimolar, mM) i blodet holdes stabilt. Glukose er et reduserende sukker, gir osmotisk effekt og høye sukkerkonsentrasjon kan gi vekst av endogene bakterier og konsentrasjonen må holdes under nøye kontroll. Etter et karbohydratrikt måltid øker utskillelsen til blodet av insulin lagret i betacellene i bukspyttkjertelen. Insulin lagret i vesikler frigis i blodet og aktiverer opptak av glukose fra blodet. Glukose blir tatt opp av glukosetransportøren GLUT4 av myocytter i skjelett- og hjertemuskelatur og lagret som glykogen. Glukose tatt opp av adipocytter blir lagret som triacylglycerider (fett). Det er noe GLUT4 i plasmamembranen, men mesteparten av GLUT4 ligger lagret i vesikler som blir aktivert av insulin og raskt fusjonere med plasmamembranen hvor GLUT4 blir inkorporert i membranen. Rab-proteinene Rab10, Rab8A og Rab13 deltar i den insulininduserte transportstyringen av GLUT4-vesiklene fram til plasmamembranen. Vanninnholdet blir regulert av vannkanalproteinet akvaporin 2 (AQP2) i epitelcellene i oppsamlingsrørene i nyrene aktivert av peptidhormonet vasopressin (antidiuretisk hormon) som gjør at mer vann blir reabsorbert fra nyrene. Signalveien fra binding av vasopressin til reseptoren går via adenylat syklase som lager syklisk AMP (cAMP) som deretter aktiver cAMP-avhengig proteinkinase A som fosforylerer AQP2. Mutasjon i genet AQP2 gir feil i oppkonsentrering av urinen styrt av vasopressin.
{kind=link}