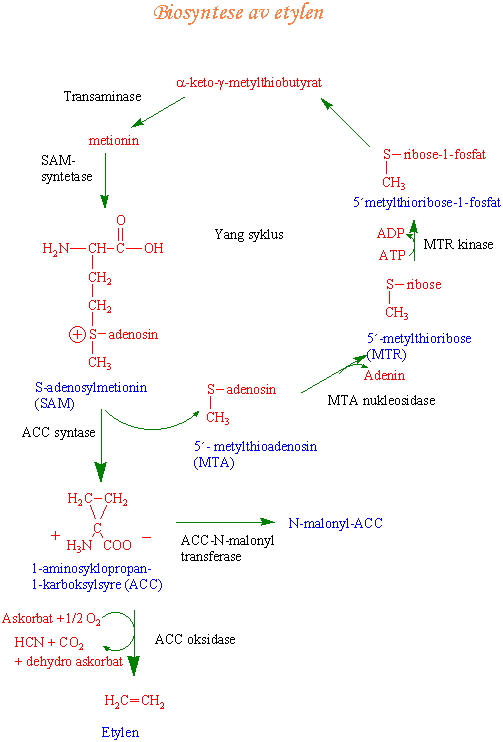
Biosyntese av etylen
Oppdagelsen av etylen og trippelresponsen
På 1800-tallet oppdaget man at kullgass til belysning gjorde at trær som vokste ved gatelamper og i nærheten av lekkasjer i ledningsnettet mistet bladene.Dimitry Neljubov ved det botaniske instituttet i St.Petersburg oppdaget i 1901 trippelresponsen på etiolerte erteplanter: redusert strekningsvekst, horisontal vekst (svelling og lateral vekst), samt agravitropisme. Dette er en tilpasning som er nyttig når frøet spirer og frøplanten skal vokse gjennom hard jord. Etylen samvirker ofte med plantehormonet auxin. Gasskromatografen med flammeionisasjonsdetektor (1959) gjorde at flere kunne gjøre undersøkelser av etylen.
Etylen - aldring og modning av frukt
Etylen induserer fruktmodning hvor konsentrasjon av organiske syrer, fenoler og tanniner synker, mens konsentrasjonen av, farge og eteriske stoffer øker. Frø klar for spredning. Nedbrytning av cellevegg og pektin gjør frukten bløt og saftig. Stivelse hydrolyseres til sukker. Karotenoider og antocyaniner produseres, noe som øker synligheten for at fruktivore dyr spiser frukten og derved sprer frøene. Moden vil si spisbar. Hvis frøene skal spres mekanisk måfruktes splittes eller spaltes.
I bananmodnerier utsettes grønne bananer for etylen som slippes ut fra gassflasker i modneriet, og derved starter banens egenproduksjon av etylen (autokatalytisk etylensyntese) og bananene blir gulfarget når grønne kloroplaster omdannes til kromoplaster med karotenoider. Når grønne tomater modner omdannes grønne kloroplaster til kromoplaster med det rødfargete karotenoidet lykopen.
Siden etylen deltar i aldring av frukt og blomster ønsker produsenter av frukt og snittblomster å redusere virkningen av etylen. Først viste man at frukt holdt seg bedre i kjølige lagringsrom med lavt trykk, lav konsentrasjon av oksygen og høy konsentrasjon av karbondioksid, jfr.
Pasteur-punkt. Man oppdaget at effekten ikke bare skyldtes effekten på respirasjonen, men at CO
2 og O
2 direkte påvirket effekten av etylen.
Etylen og holdbarhetsmidler
Etter hvert fant man at sølvioner (Ag
+) kunne hindre virkningen av etylen, og dette kan brukes som holdbarhetsmiddel i form av sølvthiosulfat på snittblomster. Etylenreseptoren inneholder kobber (Cu
2+), og man fant at flere stoffer kan binde seg til reseptoren og blokkere den, blant annet gassen 1-metylsyklopropen (1-MCP, 1-metylcyclopropene) (SmartFresh®, EthylBloc®) som bl.a. benyttes på epler, pærer og annen
klimakterisk frukt for å forsinke modningsprosessen. 1-MCP blir også brukt til å forsinke aldringen av snittblomster. Siden 1-MCP er en gass som binder seg til etylenreseptoren blir den brukt i lukkete lagerrom. Noen epler blir vokset med naturlig carnaubavoks som glansemiddel, noe som redusere transpirasjonen. Carnaubavoks (brasilvoks) blir isolert fra epidermis på bladene til den tørketålende palmen
Copernicia prunifera, syn
Copernicia cerifera og består av en kompleks blanding alifatiske voksestere, hydroksyestere, cinamatestere, syrer, alkoholer og harpikser. Skjellakk utskilt fra hunninsektet lakkskjoldlus (
Kerria lacca) som lever på blant annet kusumtreet (
Schleichera oleosa). Ingen av de godkjente voks og glansemidler skal være helseskadelige. Naturlig voks fra epler inneholder et pentasyklisk
triterpenoid (ursolsyre).

Bruk av SmartFresh. Faksimile Aftenposten Aftenposten 06.01.2006
Klimakterisk og ikke-klimakterisk frukt
Eksempler på klimakterisk frukt som produserer mye etylen samtidig med økt respirasjon under modningsprosessen er eple, tomat, banan, plomme, fersken, pære, fiken, avokado, cherimoya, mango, oliven, persimmon, og cantaloupe. Eksempler på ikke-klimakterisk frukt er citrus-frukter, vindrue, ananas, jordbær, vannmelon og kirsebær. Tomatmutanten never-ripe har en mutasjon i genet som koder for etylenreseptoren.
Sprøytemiddel som avgir etylen
Sprøytemiddelet 2-kloroetylfosfonsyre (Ethrel®, Etephon) frigir etylen i kontakt med plantevev og gir avkastning i frukt (frukttynning), gir modning av eple og tomat , og til induksjon av blomstring i ananas.
Etylen og molekylærbiologi
Arabidopsis mutanten etr1 (etylenresistent 1) har mutasjon i genet for etylenreseptorprotein, og ein2 (etylen-insensitiv 2) har en mutasjon lenger ut i signaloverføringsveien. Etylenresistente eller etyleninsensitive mutanter har etylenrespons i fravær av etylen og er en konstitutiv mutant. Etylenreseptoren ETR1 ligner bakteriell tokomponent histidin kinase, og både cytokinin og fytokrom virker etter samme prinsipielle signalvei. ETR1, ERS1, ERS2 og EIN4 er etylenreseptorer. ETR1 er lokalisert til endoplasmatisk retikulum. Etylen er hydrofob og går gjennom membraner. Etylenreseptoren trenger kobber eller zink. RAN1 (RESPONSIVE TO ANTAGONIST 1) i Arabidopsis koder for et protein som bringer kobber inn i et jerntransportprotein. Mutanten ran1 lager ikke etylenreseptor. Etylenreseptorene er aktive i fravær av etylen og etylen skrur av reseptorene.
Det er etylenresponselementer (ERE) i etylenregulerte gener. ERF1 (etylenresponsfaktor 1) koder for protein som hører med til ERE-bindende protein transkripsjonsfaktorer. EIN3 transkripsjonsfaktorer deltar i signalveien. Signalrekkefølgen er ETR1 → EIN2 → EIN3 → CTR1 →→
CTR1 ligner RAF-1, en MAPKKK serin/threonin protein kinase (mitogenaktivert protein kinase kinase kinase). To F-boks protein EBF1 og EBF2 (EIN3 binding F-boks 1 og 2) aktiverer ubiquitinering og gjør et EIN3 degraderes via proteosom. Etylen hemmer EBF/EBF2-avhengig degradering av EIN3.
Tilbake til hovedside
{kind=link}
