Nerveceller kommuniserer med andre nerveceller eller muskelceller i en kobling kalt synapser. Nevtrotransmittorer (nevrotransmittere) slippes ut av vesikler på overflaten til synapser på presynaptisk celle og diffunderer over en kort avstand i synapsekløften før de binder seg til reseptorer på neste celle (postsynaptisk celle). Nervecellene (nevroner, neuroner) overfører informasjon i kroppen hos alle dyr. Kommunikasjonen kan skje via elektriske signaler i form av et eller flere aksjonspotensialer som resultat av midlertidig åpning og lukking av ionekanaler, men kommuniseringen kan også skje via kjemiske stoffer som nevrotransmittorer. Nerveceller er gruppert i klynger kalt ganglier eller i en mer eller mindre kompleks hjerne. En nervecelle består av en cellekropp som inneholder en cellekjerne, og fra cellekroppen er det greinete utstikkere, dendritter (gr. dendron – tre) som kan motta signaler fra andre nerveceller, samt en lenger utløper, akson, kan ha lengde mer enn en meter, ogsom i enden har en greinet terminal struktur som ender i synapser som overfører nervesignalet til neste celle. Store aksoner gir mindre motstand mot aksjonspotensialet slik som hos blekksprut, artropoder og mollusker, hos blekkstronet kan aksonet ha en diameter på en millimeter. Vertebratene har trangere aksoner med liten diameter. Nevroner hos vertebrater er omgitt av gliaceller eller glia (gr. glia – lim) som beskytter aksonet. Aksonet er elektrisk isolert med en myelinskjede, med jevne mellomrom avbrutt av Ranviers knuter (noder). Myelinskjeden blir lavet av Schwannceller i det perifere nervesystemet og av glia i oligodendrocyter i sentralnervesystemet. For eksempel kan det være spenningsregulerte natrium-ionekanaler i nodiene og aksjonspotensialet kan hoppe fra node til node, saltatorisk ledning, som gir raskere overføring av aksjonspotensialet gjennom aksonet.
Aksjonspotensialet depolariserer cellemembranen i synapsen, som resulterer i åpning av spenningsregulerte kalsium-kationkanaler som gir innstrømning kalsium (Ca2+) . Økt kalsiumkonsentrasjon virker som et signal som får synapsevesiklene med nevrotransmittorer til å fusjonere med synapsemembranen og slippe fri nevrotransmittorene ut i synapsekløften. Nevrotransmittorene binder seg til ligandstyrte ionekanaler på mottakercellen (postsynaptisk celle).
Nervesignalet avsluttes 1) enten ved at et enzym bryter ned neurotransmittoren i den synaptiske kløften, for eksempel acetylkolinesterase bryter ned nevrotransmittoren acetylkolin. Eller 2) nevrotransmittoren blir tatt opp igjen i synapsen på det presynaptiske nevronet og på nytt pakket inn i vesikler. Såkalte ”lykkepiller”, som over tid fremkaller alt annet enn lykke, kan være av type serotonin reopptakshemmere, og som blokkerer for reopptaket av nevrotransmittoren serotonin som er skilt ut i synapsekløften. 3) Nevrotransmittoren kan også bindes til G-proteinkoplete som åpner eller lukker metabotrope ionekanaler. De kalles metabotrope siden de er et resultat av metabolisme i signalveien via G-proteiner. For eksempel kan hormonet noradrenalin via G-proteinkoblete reseptorer, G-protein, aktivert adenylyl syklase som gir syklisk AMP, og signalet fortsetter videre via protein kinase A som fosforylerer en ionekanal. G-protein metabotrope ionekanaler gir mer langvarig effekt enn et signal via ionotrope kanaler.
Eksempler på nevrotransmittere er:
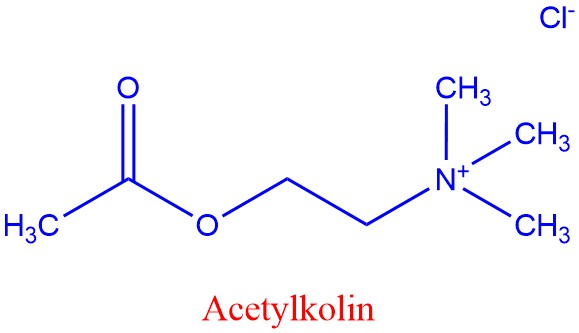
Acetylkolin
Acetylkolin som deltar i sammentrekning av muskler og kroppsbevegelse, men deltar også i hukommelse og læring. Det er to hovedtyper acetylkolinreseptorer: 1) Ligandregulerte ionekanaler som gir et eksitatorisk postsynaptisk potensial (EPSP) og 2) Nevromuskulære reseptorer som kobler en motornerve til en muskelcelle. Binding av acetylkolin til G-proteinkoblete reseptorer kan hemme enzymet acenylyl syklase (lager syklisk AMP), noe som resulterer i åpning av metabotrope kalium-ionekanaler i membranen i muskelcellen. Alkaloidet nikotin fra planter binder seg til ionotrope acetylkolinreseptorer. Botulinumtoksin hemmer frigivelse av acetylkolin.
Aminosyrer som nevrotransmittorer
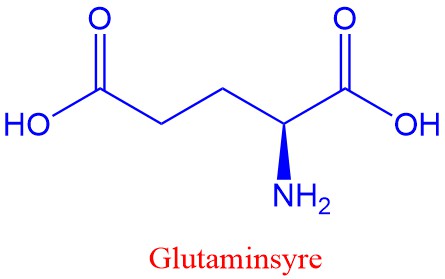
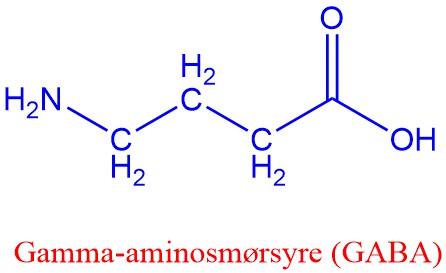
Aminosyrene glutamat (glutaminsyre), gamma-aminosmørsyre (GABA) og glysin virker som nevrotransmittorer. Hos vertebrater brukes glutamat i langtidshukommelsen i sentralnervesystemet. Imidlertid kan glutamat ikke krysse blod-hjernebarriæren, men det er allikevel noen som er skeptisk til utstrakt brukt av glutamat som smaksforsterker (umami). Hos invertebrater brukes glutamat i stedet for acetylkolin i muskelkontraksjoner.
Både GABA og glycin er hemmende nevrotransmittorer via hemmende synapsekoblinger. Binding av GABA til postsynaptisk nevron øker permeabilitet til klorid (Cl-) i anionkanaler som resulterer li et inhibitorisk postsynaptisk potensial (IPSP). Valium binder seg til GABA-reseptoren.
Biogene aminer
Serotonin, dopamin og noradrenalin er eksemper på biogene aminer laget fra aromatiske aminosyrer. Tyrosin omdannes til dopamin, og tryptofan omdannes til serotonin. Serotonin påvirker søvn, læring, hukommelse og oppstemthet.
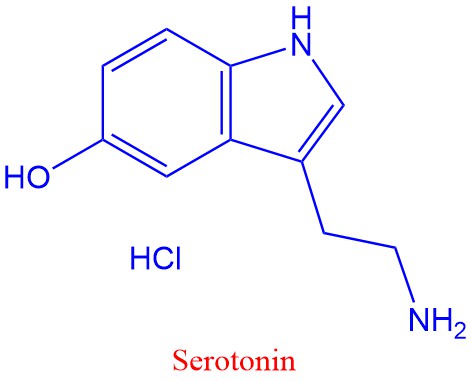
Parkinsons sykdom skyldes mangel på dopamin i hjernen. De aromatiske aminosyrene hører med til de essensielle som bare kan lages av planter og bakterier, og som vi må ha tilført via maten.
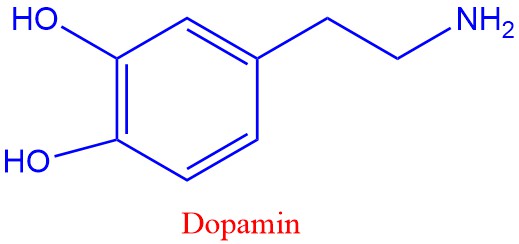
Den amerikanske immunologen Mahlon DeLong og medarbeidere arbeider med beskrive feil i de integrerte kretsene i hjernen som resulterer i Parkinsons sykdom De undersøkte en Parkinsonslignende sykdom på aper og oppdaget nevroner i en type basalganglier (subtalamisk kjerne) ble kontinuerlig avfyrt og ved å blokkere disse ble skjelvingene redusert, enten ved å fjerne nevronene eller foreta dyp hjernestimulering med høyfrekvent strøm av dem. Mahlon DeLong fikk Breakthrough-prisen i livsvitenskap i 2014. Skjelvingene skyldes minsket evne til å produsere dopamin i basalgangliene.
Michel Houellebecq: Sérotonine (2019)
Ionekanaler nevrotransmittere
Reseptorer for nevrotransmittoren acetylkolin er ligandstyrte ionekanaler. Andre ligandstyrte reseptorionekanaler som responderer på ekstracellulære signaler er GABA (gamma-aminosmørsyre) reseptorkanal spesifikk for anionet klorid (Cl-) som binder nevrotransmittoren GABA. Glycinreseptorionekaner binder nevrotransmitteren glycin og er spesifikke for hydrogenkarbonat (HCO3-).
Serotonin reseptorionekanal som binder nevrotransmittoren serotonin er en kationkanal. Andre ligandregulerte ionekanaler bruker den ekstracellulære ligander er syklisk GMP (3’-5’-syklisk guanosin monofosfat, cGMP). Andre ligander i ligandstyrte ionekanaler er syklisk AMP (3’-5’-syklisk adenosin monofosfat, cAMP). IP3 (inositol-1,4-5 trifosfat) og adenosintrifosfat (ATP).
Gassene NO og CO som nevrotransmittorer
Nitrogenmonoksid (NO) er et radikal laget fra aminosyren arginin hos mennesker, og som deltar i signalveien for avslapning av muskelatur, jfr. virkningen av potensmiddelet Viagra. Karbonmonoksid (CO) er ekstremt giftig når det blokkerer cellerespirasjonen i mitokondriene, men det har vist seg at små mengder CO kan ha en funksjon som nevrotransmittor.
Nevropeptider
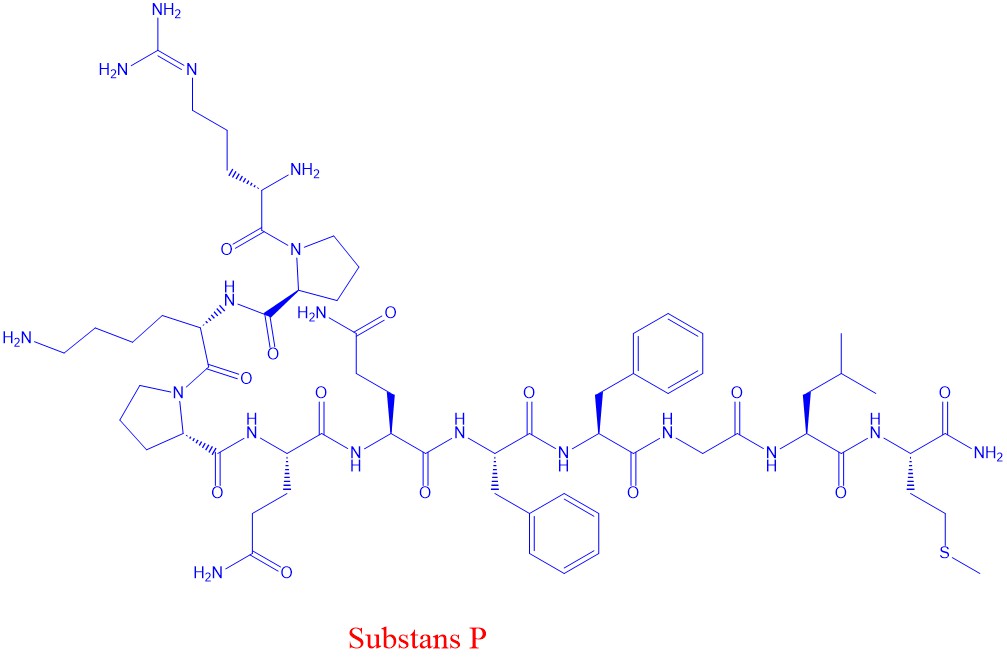
Substand P og metenkefalin (et endorfin) er eksempler på peptider som virker som nevrotransmittorer. Endorfiner minsker smerte og kan gi euforia. Opiater fra opiumsvalumen virker ved å binde seg til opiatreseptorer, som også binder endorfiner.