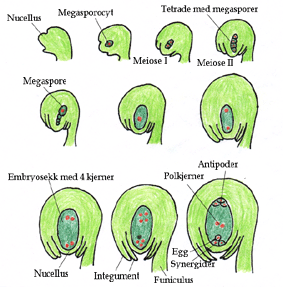
En arkesporcelle i cellelaget under epidermis i nucellus i tidlig stadium av utvikling av frøemnet i fruktknuten får en stor cellekjerne, og danner en megasporocyt.
Dannelse av embryosekk i frøemne hos dekkfrøete planter (angiospermer)
Megesporocyten fungerer som megasporemorcelle. Megasporemorcellen gjennomgår to reduksjonsdelinger, meiose I og II, hvor det blir dannet fire haploide megasporer i makrosporogenese. I dette stadiet dannes anlegg for integumentene i nucellus, og integumentene vokser i retning mot chalaza etter hvert som embryosekken øker i størrelse. Nucellus virker som næringsvev for embryosekken, men hos mange arter forsvinner nucellus når integumentene utvikler seg. Det indre integumentet nærmest embryosekken utvikler seg i stedet til et cellelag kalt endotelium, og som har samme funksjon som tapetlaget i pollenknappene hos den hannlige gametofytten, og gir næring til embryosekken.
Hos monosporiske arter dør tre av megasporene nærmest mikropylen i kontrollert programmert celledød, mens den fjerde makrosporen nærmest chalaza gjennomgår tre mitoser (mitose I, II og III) i en makrosporegametogenese hvor det blir en flerkjernet celle med 8 kjerner i et syncytium uten cytokinese. Denne fjerde makrosporen inneholder plastider, mitokondrier i et tett cytoplasma og gir maternal nedarving av disse organellene med prokaryot opprinnelse. Etter mitose I er det en kjerne i hver ende av embryosekken, og det etableres en polaritet i aksen mikropyle til chalaza. I mitose II er de to kjerner i hver ende, og i mitose II beveger en kjerne fra hver ende seg mot midten og danner sentralkjerner i sentralcellen. Nærmest mikropylen cellulariseres eggcellen og to synergider på hver side av eggcellen, atskilt med mer eller mindre tykke cellevegger og danner et eggapparatet. Synergidene har et filiformapparat med utvekster fra celleveggene som går ut i nucellus. I chalazaenden cellulariseres tre antipodeceller. Tto polkjerner fra hver sin ende av embryosekken (mikropyleenden og chalazaenden) beveger seg til sentrum av den store sentralcellen med stor vakuole. Dette er den vanligste formen for embryosekk, Polygonum-typen.
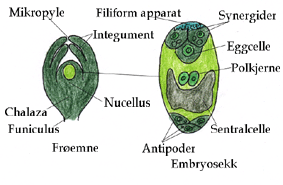
8-kjernes embryosekk hos en dekkfrøet plante
Løk (Allium) er bisporisk, det vil si to celler fra tetraden danner en makrospore med to cellekjerner, etterfulgt av to mitoser gir en vanlig Polygonium embryosekk. Moskusurt (Adoxa) i moskusurtfamilien har en tetrasporisk embryosekk hvor alle de fire cellene i tetraden danner en megaspore med fire cellekjerner, etterfulgt av en mitose.
Når spissen av pollenslangen brister og frigjør de to spermcellene fusjonerer den ene spermcellene med eggecellen og to polkjerner fusjonerer med denandre spermcellen og lager triploid endosperm. I slekten Peperomia er det 8 polkjerner og det dannes 15n endosperm.
Diploid endosperm
I nøkkerosefamilien (Nymphaceae) og mjølkefamilien (Onagraceae) er det en firekjernet embryosekk (hunnlig gametofytt) uten antipoder. Fusjon med den ene av de to spermkjernene fra pollenslangen danner en primær diploid endospermkjerne. Hos noen degenerer antipodene før befruktning. I nattlysslekten Oenothera er det også bare en polkjerne slik at det dannes diploid endosperm. Oenothera er også monosporisk, men det er den cellen i tetraden som er nærmest mikropylen som overlever, mens de tre nærmest chalaza desintegrerer. Det er bare to vekstdelinger (mitoser) slik at embryosekken inneholder bare fire cellekjerner, fordelt på en eggcelle med to synergideceller nærmest mikropylen, og en sentralcelle med en cellekjerne. Diploid endosperm er en basal evolusjonær karakter.
I utviklingen av blomsterplantene danner Amborella trichopoda, og arter i orden vannliljer (Nympheales), samt i Austrobaileyaceae med slekten Austrobaileya endemisk i Australia, stjerneanisfamilien (Illiciaceae) med slekten stjerneanis (Illicium), kadsurafamilien (Schisandraceae), samt familien Trimeniaceae en evolusjonær tidlig, opprinnelig og basal utviklingslinje. Slike arter har primitive plesiomorfe karakterer i blomst (mange blomsterblad i spiralplassering (mange pollenblad, oversittende fruktknute med mange fruktblad), radiærsymmetrisk blomst, diploid biparental endosperm) og primitivt ledningsvev. Amborella trichopoda som vokser endemisk på Ny-Caledonia, danner en søstergruppe til blomsterplantene (angiospermene), har ledningsvev med bare trakeider og ikke vedrør for transport av vann og mineralnæring, slik som gymnospermene og karsporeplantene. Denne utviklingslinjen ble etablert før de enfrøbladete og tofrøbladete plantene.
Apomiksis
Ved apomiksis utvikles den hunnlige gametofytten og embryosekken uten at det har skjedd en reduksjonsdeling (meiose). Det vil si at embryosekken dannes fra en allerede diploid celle i frøemnet.
Embryogenese
Den ene generative cellen i pollenlangen fusjonerer med de to polkjernene, mens den andre generative cellen med spermkjernen fusjonerer med eggapparatet. Spermkjernen fusjonerer med eggcellen og danner en diploid zygote, starten på embryogenesen. Allerede ved første asymmetriske celledeling av zygoten med en stor basalcelle og en mindre apikalcelle etableres en polaritet i det som skal utvikle seg til et embryo. Det skjer en reorganisering av cytoskjelettet i apikalcellen som deler seg to ganger på langs (longitudinelt) og to ganger på tvers (transversalt) og danner et proembryo bestående av åtte celler (oktantstadiet). Basalcellen deler seg på tvers gjentatte ganger og danner hypofysis og suspensor. Påfølgende celledelinger i proembryo gir apikale meristemer i rot- og skuddanlegg, frøblad, og hypokotyle.
Hjertestadiet hos tofrøbladete med et apikalt skuddmeristem mellom de to frøbladene. Bilateral symmetri hos tofrøbladete kan observeres i hjertestadiet med provaskulært vev, etterfulgt av torpedostadiet og frøbladstadiet. Embryoskuddaksen består av embryorot (kimrot, radikel), hypokotyle, epikotyle og en skuddapikal knopp (kimknopp, plumula). Hos de enfrøbladete er det ene frøbladet dekket av en beskyttende koleoptile og kimrota er dekket av en koleorhiza.
Den beskyttende fruktknuten (ovariet) vokser og blir til frukt.
Den øverste cellen i suspensor nærmest embryo, hypofysis, blir den del av kolumella i rothetta. Seinere i embryoutvikling dør suspensor ved programmert kontrollert celledød.