Den moderne arvelæren startet i 1866 med at den østeriske munken Gregor Mendel publiserte kryssingsforsøk med erter dyrket i urtehagen - Versuche über Pflanzen-Hybriden. Valget av hageerter som forsøksmateriale var meget gunstig.

Erter er ettårige, og med lukkete og tokjønnete blomster er de selvpollinerende og selvfertile.

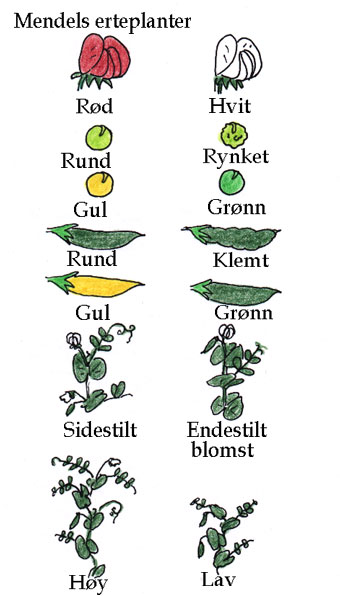
Erter (Pisum sativum) med blomster og belger.
Erter kan i tillegg krysspollineres kunstig ved å overføre pollen fra en plante til arrene i blomstene på en annen plante, hvor pollenbærerne først er fjernet før de er ferdig utviklet. Det fantes mange konstante varieteter av erter, og de karakterene Mendel undersøkte var belgenes og ertenes form og farge, blomsterfargen, blomstenes plassering endestilt eller aksialt, og høyden på plantene. Enten-eller egenskapene Mendel studerte systematisk var lokalisert på 7 (n) kromosomer. Han observerte at høye erteplanter fikk frø som ga like høye erteplanter når ertene ble sådd på nytt. En linje erter laget alltid runde erter, en annen linje laget alltid rynkete erter. Dette er fenotype karakteristikker. Mendels eksperimenter viste at fenotyp karakteristikkene ble bestemt av arvlige faktorer, seinere kalt gener, som ble overført med kjønnscellene til neste generasjon. Mendel lurte på hva som ville skje hvis han kunstig krysset en kort erteplante med en lang. Kryssing mellom to varieteter med forskjellige karakterer kalles hybridisering. Hybridene ble ingen mellomting mellom kort og lang. Ertene ble tatt ut av belgene og sådd på nytt. Det viste seg at ertene fra hybridene bare ga høye planter. Derimot, i neste generasjon (F2-generasjonen), da plantene ble naturlig selvpollinert, og ertene fra disse plantene ble sådd på nytt, ga disse ca. 3/4 med lange erteplanter og 1/4 med korte erteplanter. Mendel var overbevist om at avkommet etter første kryssing inneholdt enheter for egenskapen korte erteplanter, selv om alle var lange, og at disse enhetene kunne nedarves. Lignende forsøk gjorde Mendel med grønne, gule, rynkete eller glatte erter, samt purpurfargede eller hvite blomster.

Vi kan idag forklare Mendels resultater ut fra kunnskapen om dominante og recessive alleler (genutgaver) og genenes plassering på kromosomer. Et gen kan ha forskjellig uttrykk av en karakter (alleler). En diploid organisme vil ha to alleler for hvert gen, ett på hver av de homologe kromosomparene i en diploid organisme. Et allel som uttrykkes i fenotypen i F1- generasjonen (filialgenerasjonen, avkom) kalles dominant. Et allel som ikke uttrykkes kalles recessivt. Fenotypen er et resultat av virkningen av mange gener. Mendel forsøkte å finne grunnlaget for hvordan utseendet av planten (fenotypen) kunne forklares ved arvelige faktorer (genotypen). En monohybrid kryssing vil si at foreldrene (parentalgenerasjonen, P- generasjonen) atskiller seg bare i en genetisk egenskap f.eks. gul og grønn farge på ertene. Gul farge er dominant over grønn. Kryssing mellom konstante gule erter (GG) med konstante grønne (gg) vil gi planter med kjønnsceller som henholdsvis enten inneholder allelene G eller g. Mendel innførte bruken av stor bokstav for et dominant arveanlegg og liten bokstav for et recessivt arveanlegg. Alle zygoter etter denne kryssingen, F1-generasjonen, vil være av formen Gg. En gul fenotyp har en genotyp med allelkombinasjon GG eller Gg. Selvpollinering blant F1- generasjonen vil gi en F2-generasjon med gule og grønne erter i forholdet 3:1. En plante er homozygot for et gen hvis begge allelene er like, f.eks. formen GG eller gg. Planten er heterozygot for et gen hvis genet har to forskjellige alleler (Gg). Arveanleggene fordeles likt på hannlige og hunnlige kjønnsceller. En kryssing mellom planter med runde erter (RR) med planter med rynkete erter (rr) gir en F1-generasjon med bare runde erter (Rr). Karakteren runde erter er dominant over rynkete erter. Plantene i F1-generasjonen kan ved selvbefruktning gi andre filial generasjonen (F2), og nå dukker runde og rynkete erter opp i forholdet 3:1, som er forholdstallet ved en monohybrid kryssing.
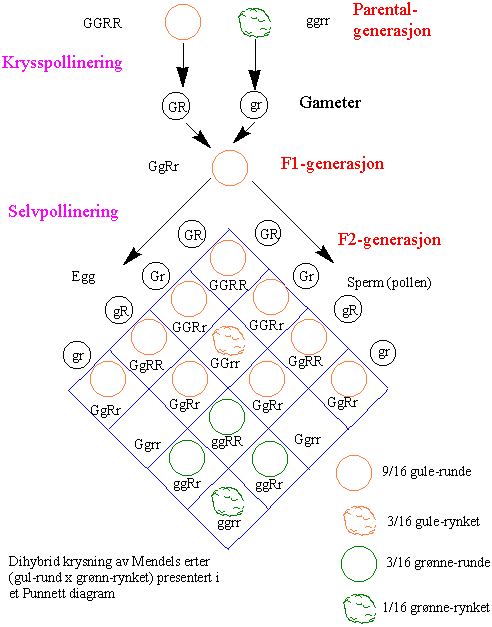
I en dihybrid kryssing vil foreldrene atskille seg i to karakterer med to par med gener lokalisert på forskjellige kromosomer f. eks. karakterene grønn eller gul farge, og runde eller rynkete erter. Det finnes trihybrid kryssing o.s.v.
{kind=link}
Forholdstallet i en dihybrid kryssing er 9:3:3:1.
Vi vet i utgangspunktet ingenting om genotypen ut fra fenotypen. I mange tilfeller er ingen av genene i et par dominante. Krysses f.eks. en varietet av løvemunn med rød blomster med en varietet med hvite blomster får F1-avkommet rosa blomster. I F2-generasjonen blir forholdet i avkommet 1 rød : 2 rosa : 1 hvit. Dette kalles ufullstendig dominans. Omgivelsene kan påvirke i hvilken grad et gen uttrykkes.
Tilbakekryssing (testkryssing) utføres ved å krysse en hybrid med en av sine foreldre, noe som gir et 1:1 forhold (monohybrid kryssing) eller 1:1:1:1 (dihybrid kryssing). F.eks. kryssing mellom F1-generasjonen og en recessiv forelder (Aa x aa) vil gi avkom med Aa og aa i forholdet 1:1. Kryssing mellom F1-generasjonen og dominant forelder (Aa x AA) vil gi dominante Aa og AA i forholdet 1:1.
Mendels 1. lov sier at det er forskjellige utgaver av et gen (alleler), og at i kjønnsceller som dannes i diploide individer vil allelene bli atskilt fra hverandre. Hver kjønnscelle har like stor mulighet til å få et av allelene. Selv om et diploid individ ikke kan ha mer en to alleler ad gangen for et gen, kan populasjonen som sådan ha en rekke multiple alleler (polymorfi). Hvert kromosom har imidlertid bare ett allel for ett gen. Mendels 2. lov sier at gener lokalisert på forskjellige kromosomer sorteres uavhengig av hverandre. Mendels lover burde egentlig kalles prinsipper siden det finnes unntak.
Hver organisme har et karakteristisk antall kromosomer. Hver diploid vegetativ celle har dobbelt sett med kromosomer, ett sett som de har fått fra moren og ett sett som de har fått fra faren. Hvert kromosom i den vegetative cellen har en homolog. Hvert par med homologe kromosomer inneholder samme genetisk informasjon, men innholdet i genene kan ha forskjellige utgaver (alleler). Kjønnscellene (spermceller og eggceller) er haploide, slik at de har bare ett sett av kromosomene. Sammensmelting av spermcelle og eggcelle ved befruktning, gjenoppretter det diploide cellestadiet som finnes i planten. Imidlertid er polyploidi vanlig i planter.
Mange gener er bundet til andre gener og arves sammen. Dette fordi hvert gen har en spesiell posisjon på kromosomet. Hvert kromosom er et langt DNA-molekyl koblet til forskjellige proteiner. Hvert gen har en spesifikk plassering på kromosomet kalt locus. Genkart kan lages ved å beregne % overkryssing Overkryssing vil skje oftere mellom to gener som ligger langt fra hverandre enn mellom to som ligger nær hverandre. Avstanden mellom gener måles som frekvens av rekombinasjon. En genkartenhet er definert som avstanden hvor man forventer overkryssing i 1 av 100 gameter. Denne enheten, 1 % rekombinasjon, kalles centimorgan etter genetikeren T. H. Morgan.
Mendel hadde ikke kunnskap om kromosomer og fordeling av disse under meiosen. Allikevel kom han fram til allmenngyldige arvelover. Mendel arbeidet også med andre planter, bl.a. sveve (Hieracium). Sveve har imidlertid apomiksis slik at Mendel klarte ikke å bekrefte funnene gjort med erter. Mendels banebrytende arbeidet ble gjenoppdaget i år 1900 av de Vries, Correns og Tschermak.
Hvorfor så Mendels erter ut som de gjorde ?
Fiolette versus hvite blomster
Rød, blå eller fiolett farge på blomster skyldes vanligvis en type flavonoider kalt anthocyaniner. Anthocyaninene lages i en lang kjede med biokjemiske reaksjoner, hvert trinn katalysert av et enzym. Hvis et gen som koder for et enzym i kjeden har en endring i basesekvensen som gjør at det ikke virker, mister planten evnen til å lage fargede anthocyaniner, og får derved hvite blomster.
Høye versus lave planter
Plantehormonet gibberellin er nødvendig for at planter skal kunne strekke seg. Gibberellin lages i en lang biosyntesevei med mange enzymatiske trinn. Hvis et gen har mistet evnen til å lage et enzym som er nødvendig for biosyntesen av gibberellin vil ikke planten lenger ha mulighet til å lage gibberellin, og blir derved kort.
Grønne og gule erter og belger
Grønnfargen skyldes klorofyll og gulfargen karotenoider. Når ertene utvikles i belgen er de grønne. Når ertene og belgen tørker ut vil klorofyll brytes ned av enzymer. Er det et ikkefungerende gen i planten som har med biosyntesen av klorofyllnedbrytende enzymer, vil ertene forbli grønne.
{kind=link}
Runde versus rynkete erter
Erter fyller frøbladene med store mengder stivelse som skal brukes som opplagsnæring for den unge kimplanten. Stivelse er satt sammen av glukose og består av amylose og amylopektin. Stivelse lages fra sukrose som kommer fra bladenes fotosyntese. En gruppe gener koder for enzymene som skal lage stivelse. Rynkete erter mangler et enzym som har med syntese av greining av stivelsesmolekylene i amylopektin. Dette skyldes at et transposon har satt seg inn i strukturgenet, og gitt et defekt recesivt allel, i homozygot form rr. Homozygote dominante planter har allelene RR. Siden de dobbeltrecessive plantene får dårligere evne til å lage stivelse fylles ertene istedet med mer sukrose. Sukrose virker, i motsetning til stivelse, meget osmotisk aktivt og tiltrekker seg vann. I det siste stadiet i frøutviklingen tørker frøet ut, og siden rr frø inneholder større mengder vann vil dette gi mer rynkete frø når de tørker, sammenlignet med erter som har normal syntese av stivelse. Et gen koder for et protein, som er et enzym som katalyserer en biokjemisk reaksjon i cellen, og dette igjen vil påvirke fenotypen. Planter som er forskjellige bare i RR og rr er helt like bortsett fra frøenes egenskaper. rr frøene har også mindre mengde av lagringsproteinet legumin og høyere fettinnhold en frø fra RR plantene. Disse forskjellene er pleiotrofe egenskaper til r allelet. En enkel forandring eller mutasjon i et gen kan påvirke flere fenotypeffekter i en plante og disse multiple fenotypeffektene sies å være pleiotrope.
Kvantitative arvbare egenskaper
Mendel undersøkte enten-eller karakterer, men mange arvelige egenskaper har flytende overganger og er kvantitaive arvbare egensaper. Kvantitative størrelser kontrolleres av flere kvantitative arvbare loci ved at disse virker sammen.
Moderne syntese
Moderne syntese innen evolusjonsbiologi er amalgamering av Darwins naturlig seleksjonsteori, Mendels genetikk og arv og populasjonsgenetikk som forklarer opprinnelsen til artene, fenotypevariasjon, mutasjoner og biologisk biodiversitet. Begrepet ble brukt av zoologen Julian S Huxley I boka Evolution: The modern synthesis (1943). Begrepet har tilknytning til utvidet evolusjonær syntese (EES) brukt av Ernst Mayr og neo-darwinisme.