Fotorespirasjon
Karbondioksid og nitrogen i form av ammonium (NH4+) frigis i mitokondriene i overgangen fra glycin til aminosyren serin. Karbondioksid utskilles og går tapt, men ammonium reassimileres via enzymsystemet GS-GOGAT.. Fotorespirasjon finnes hos C3-plantene, men tilsynelatende ikke hos C4-plantene og CAM- plantene, fordi utskilt karbondioksid blir effektivt fanget opp av enzymet fosfoenolpyruvat karboksylase som disse har. Fotorespirasjonen øker med temperaturen fordi løseligheten av CO2 i vann minsker raskere enn for oksygen ved økende temperatur. Dessuten gir økt temperatur økt Km-verdi for karboksyleringsreaksjonen, men ikke tilsvarende økning i Km-verdien for oksygenasereaksjonen.
Mer om fotorespirasjon
Oksygenutviklingen under fotosyntesen kan være hundre ganger større enn oksygenopptaket i mørke. I lys har C3-planter lysavhengig opptak av oksygen og utskillelse av CO2 koblet til syntese og metabolisme av glykolat, kalt fotorespirasjon.
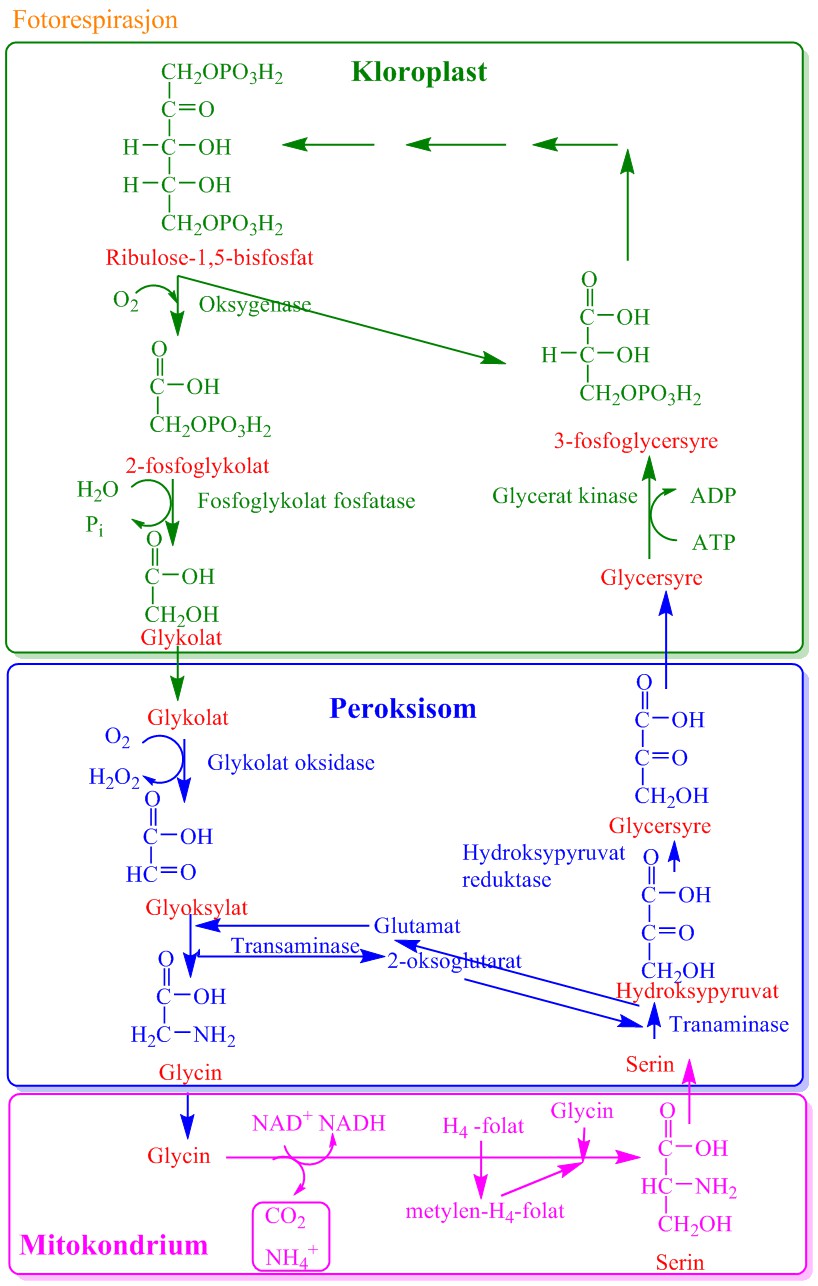
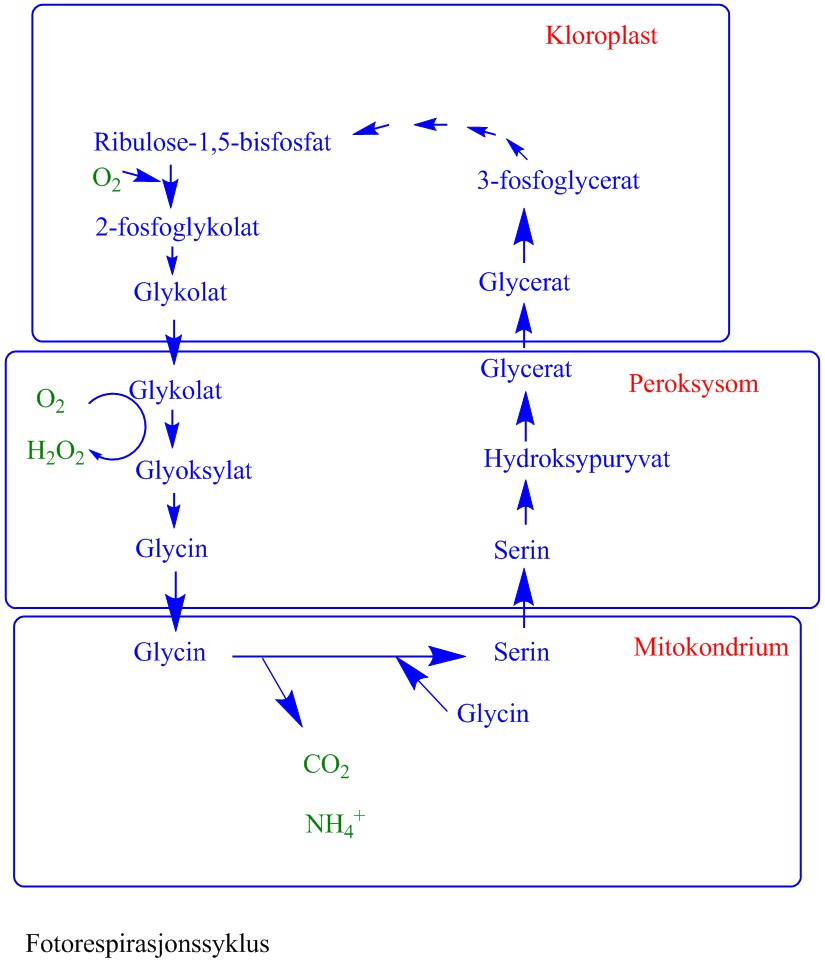
Rubisko har oksygengaseaktivitet hvor oksygen og karbondioksid konkurrerer om binding til samme aktive sete. Siden både oksygen og karbondioksid konkurrerer om binding til rubisko blir konsentrasjonen av disse gassene viktig for fotosyntesen Oksygenaseaktiviteten til rubisko gir 2-fosfoglykolat og 3-fosfoglycersyre ved binding av O2 istedet for CO2.
ribulose-1,5-bisfosfat + O2 → 2-fosfoglykolat + 3-fosfoglycersyre
Fosfoglykolat kan bli resirkulert tilbake til 3-fosfoglycersyre, men ca. 25 % tapes som CO2 i mitokondriene. I motsetning til vanlig respirasjon i mitokondriene hvor det lagres energi, forbruker fotorespirasjonen energi. Opptil 40 % av assimilert karbon i fotosyntesen kan tapes via fotorespirasjon. Fosfoglykolat er substrat i den fotorespiratoriske omsetningsveien som resulterer i tap av CO2
Fosfoglykolat hydrolyseres til glykolat katalysert av fosfoglykolat fosfatase.
2-fosfoglykolat + H2O → glykolat + Pi
Glykolat overføres til peroksysomene hvor glykolat oksideres av glykolat oksidase til glyoksylat og hydrogenperoksid (H2O2). Glykolat oksidase er et FMN-enzym (et flavoprotein:
glykolat + O2 → glyoksylat + H2O2
Dette enzymet kan også oksidere glyoksylat til oksalsyre:
glyoksylat + O2 → oksalsyre + H2O2
Noen alger bruker glykolat dehydrogenase istedet for glykolat oksidase. Hydrogenperoksid som lages av glykolat oksidase fjernes av katalase i peroksysomene. Glyoksylat omdannes til aminosyrene glycin katalysert av glutamat-glyoksylat aminotransferase.
glyoksylat + glutamat ⇔ glycin + 2-oksoglutarat
Glycin fraktes deretter til mitokondriene hvor to molekyler glycin gir ett molekyl av aminosyren serin og CO2 (fotorespirasjon) og ammonium (NH4+). Glycindekarboksyleringen er koblet til en serin hydroksymetyltransferase:
glycin + tetrahydrofolsyre + NAD+ → 5,10-metylentetrahydrfolsyre + CO2 + NH3 + NADH
Denne reaksjonen katalyseres av serin hydroksymetyltransferase og bruker metylen-tetrahydrofolinsyre som overfører av en-karbonet.
C1-tetrahydrofolinsyre kan brukes til overføring av en-karbon, men i fotorespirasjon kondenseres den til ett molekyl til med glycin:
glycin + 5,10-metylentetrahydrfolsyre → serin + tetrahydrofolsyre
Serin fraktes tilbake til peroksisomene hvor serin ved transaminering omdannes til hydroksypyruvat etterfulgt av reduksjon til glycersyre katalysert av hydroksypuruvat reduktase. Glycersyre fraktes tilbake til kloroplasten hvor det ved hjelp av ATP omdannes til 3-fosfoglycersyre katalysert av en kinase.
Ammonium (NH4+) dannet i fotorespirasjon blir reassimilert via glutamat og glutamin (GS-GOGAT) i en fotorespiratorisk nitrogensyklus.
Ved økt temperatur er CO2 mindre løselig enn oksygen slik at forholdet forrykkes med økt fotorespirasjon som resultat.
Hvorfor plantene har fotorespirasjonen har er omdiskutert. Siden også anaerobe fotosyntetiske bakterier har oksygenaseaktivitet i rubisko kan det er evolusjonære relikter som i utgangspunktet var tilpasset en lavere oksygenkonsentrasjon. Det mest sannsynlige er at fotorespirasjonen beskytter plantene mot fotooksidasjon og fotoinhibering ved å senke oksygenkonsentrasjonen. Fotorespirasjon fjerner ATP, reduksjonskraft og oksygen.
Økende temperatur påvirker løseligheten av CO2 og ved økende temperatur minker konsentrasjonen av CO2 mer enn O2, noe som gir økt fotorespirasjon ved økende temperatur.
CO2-anrikning ved rubisko
C4-planter, CAM-planter, alger og blågrønnbakterier kan oppkonsentrere CO2 i nærheten av rubisko og på denne måten unngå fotorespirasjon. Aktiv innpumping av bikarbonat (HCO3-) og deretter overføring av bikarbonat til CO2 katalysert av karbon anhydrase kan gi anrikning av karbondioksid og derav lavere fotorespirasjon.
Mange alger har ikke fotorespirasjon grunnet oppkonsentrering av CO2. CO2-pumpen i alger er en gasskanal som er homolog med Rheus protein i erytrocytter i blodet.
CO2 og HCO3- pumper i plasmamembranen ved lav konsentrasjon av CO2 er assosiert med NAD(P)H dehydrogenase. I cyanobakterier fraktes hydrogenkarbonat i cytosol til karboksysomer bestående av rubisko omgitt av et proteinskall.