Virus finnes i enormt antall på Jorden der hvor det er levende liv og celler. Bakteriofagene hindrer uhemmet økning i bakteriepopulasjoner på land og i vann. I havet har man funnet at de fleste av bakteriene er infektert med bakteriofager. Siden definisjonen av "liv" er koblet til en celle, blir virus ikke betraktet som liv, men som cellulære parasitter. Bakteriofagene inngår i rustningskappløpet, krigføringen og konkurransen mellom bakterier, aktinobakterier (tidliger ekalt strålesop (aktinomyceter) , samt sopp, for eksempel i en komposthaug med mye organisk materiale, et rustningskappløp forklart av Rød dronning hypotesen.
Enterobakteria fag lambda (fag λ) i orden Caudovirales er en temperat fag som kan enten formere seg i lytisk syklus eller lysogen syklus og inneholder et lineært dobbelttrådet DNA (48490 basepar). fag λ består av et hode med kapsid formet som et ikosaeder og inneholder DNA, og en hale som ender i halefibre (haletipp) som den bruker til å feste seg til bakterieoverflaten.
Bakteriofager kan infektere kan være spesifikk for en bakterie eller infekterflere bakterieslekter. Temperat fag P2 i virusslekten Peduovirus, med dobbelttrådet DNA pakket i et ikosaederkapsid kan infektere tarmbakterien Escherichia colii, men også Serratia, Yersiinia, og Klebsielle pneumoniae. Faggenomet injiseres i bakterien, men ikosaederkapsidet forblir på utsiden festet med proteinhalefibre til bakterieoverflaten. En temperat fag vil si at den kan formeres direkte, replikeres med bakteriens ribosomerog lager proteiner for kapsid og faghale, gir lysis (bakterien spekker og slipper fri nye bakteriofagparikler (lytisk syklus) og har med egne lysisgener, eller eller den kan gå inn i lysogen syklus ved å fusjonere og inkorporere faggenomet med bakteriegenomet. Enterobakteri a fag P4 i E.coli har et dobbelttrådet lineært DNA-genom og virker som et satelittvirus og kan bare gå inni lytisk syklus i samvirke med fag P2, men kan selv gå inn i lysogen syklus og genomet kan også formere seg som et plasmid i bakteriecellen.
Enterobakteria fag T2 er et bakterievirus i orden Caudovirales, slekt Tequatrovirus med dobbelttrådet DNA som infekterer E. coli. Fag T2, T4 og T6 kalles T-liketallsfag. Bakteriofag T7 infekterer også E. coli og går inn i en lytisk syklus. Genomet til T7 ble sekvensert i 1983 bestående av et 40 kbp dobbelttrådet DNA som koder for ca. 55 proteiner. Fag T1-T7 er T-oddetalsfager. Myovirus i familien Myoviridae i orden Caudovirales kan infektere ekte bakterier eller arkebakterier, T4 er et myovirus.
Fag phiX174 ( φX174) har et (+) sens sirkulært enkelttrådet DNA genom (5386 nukleotider), koder for 11 proteiner og kan infektere E. coli. φX174 genomet har blitt klonet i gjær og inngår i syntetisk biologi.
Inovirus i familien Inoviridae inneholder filamentformete bakteriofager e.g. fag M13 i E. coli med et enkelttrådet sirkulært DNA, og noen av dem kan inngå i biofilm. Fag M13 er en kronisk fag som ikke dreper bakteriecellene. M13 plasmider er anvendt innen rekombinant DNA-teknikker og fag "display".
Bakteriofagene fester seg til veggen til bakteriene, og sender faggenomet inn i cellene via injeksjon. Genomet til fagen kan bli inkorporert i genomet til bakterien og danner en profag som del av en lysogen syklus, eller som et episom. Inaktive profag plassert inn i bakteriecellens DNA blir replikert ved celledelingen i bakteriene, og kan holde seg inaktive i tusenvis av bakteriegenerasjoner. Alternativt er en lytisk syklus hvor bakterien produserer fagpartikler for deretter brister og dør i en lytisk syklus.Virusgenomet bruker bakteriens RNA polymerase (eller DAN-polymerase) og skrur av bakteriens gentranskripsjon. En nuklease kodet av bakteriofaggenomet kutter opp bakteriekromosomet og gir nukleotider som trengs for å lage bakteriofaggenom. Andre gener fra bakteriofagen lager kapsidprotein, samt et enzym som gir lysis og frigir bakteriofagpartiklene. Prosessen fra infeksjon til lysis kan være utført i løpet av ca. 30 minutter. I en lytisk syklus vil bakterien danne en plakk, en klar sone på et agarmedium i en petriskål.
Noen bakteriofager kan brukes som kloningsvektor, bl.a. bakteriofag lambda.
Bakteriene har utviklet en rekke strategier for å bekjempe bakteriofager og fremmed DNA, bl.a. restriksjonsenzymer og CRISPR-Cas9.
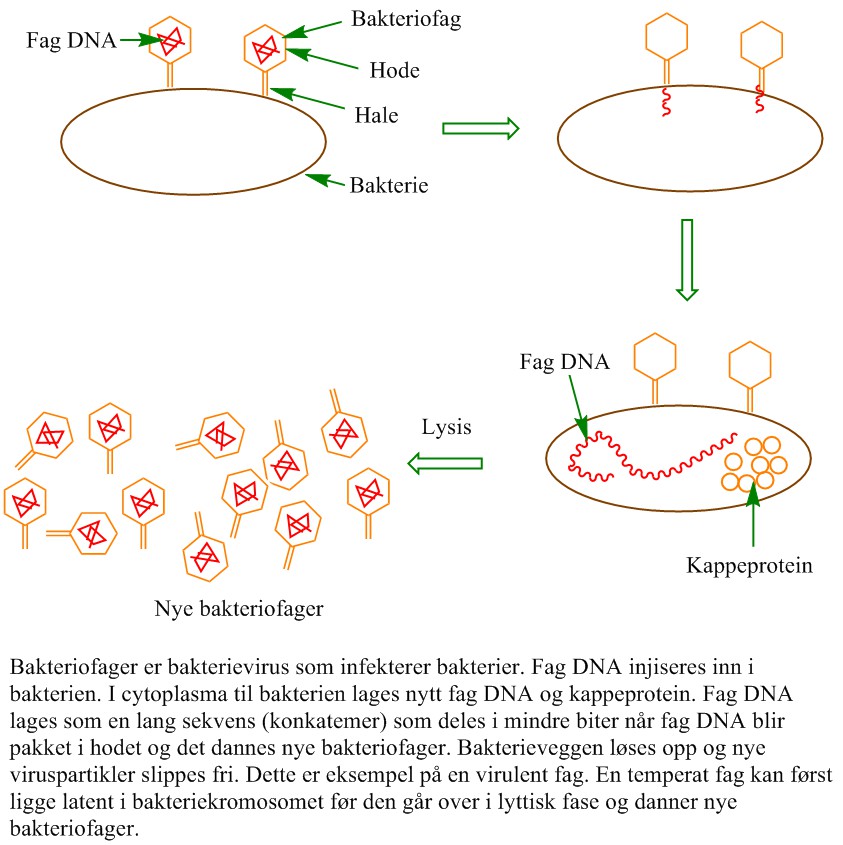
Bakteriofager er komplekse enkelttrådete eller dobbelttrådet DNA-virus som angriper bakterier. Bakteriofager har et genom med DNA eller RNA innkapslet i et proteinskall. Etter at genomet er injisert blir det replikert cytoplasma i bakterien.Fibre på halen til bakteriofagen fester viruset til bakterien. Bakteriofager holder bestanden av bakterier i sjakk og spiller derfor stor rolle i Jordens økosystemer.
ICTV (International Committee on Taxonomy of Viruses) har laget et taksonomisk system for bakteriofager som infekterer bakterier og arkebakterier, basert på morfologi og type nukleinsyrer (lineær dobbelttrådet DNA (dsDNA), sirkulær dsDNA, sirkulær enkelttrådet DNA (ssDNA), lineær enkelttrådet RNA (ssRNA), segmentert dobbelttrådet RNA (dsRNA).
Bakteriofager kan reprodusere seg i enten en lytisk syklus eller lysogen syklus. I den lytiske syklus dannes bakteriofagene og vertsbakterien dør (lysis). Lysogene bakteriofager setter DNA inn i vertscellen ved rekombinasjon. I en lysogen syklus replikeres fagen i vertsgenomet uten å ødelegge celler. Lysogene celler er resistente mot ny infeksjon av en tilsvarende fag.
Temperate fag, for eksempel fag lambda benytter både lytisk og lysogen syklus. Antisens-RNA deltar i valget mellom disse to fasene. Fag lambda fester seg på utsiden av bakterien Escherichia coli, og sender inn fag lamda-DNA inn i vertscellen. Inne i bakterien danner lamda DNA en ring og går enten inn i lytisk eller lysogen syklus. I lysogen syklus plasseres fag lamda-DNA inn i vertscellens DNA ved rekombinasjon og blir et til profag.
Et integrert bakteriofag i bakterecellens DNA kalles en profag. Profagen replikeres sammen med bakteriens DNA. Hver gang E.coli deler seg så blir fag lamda DNA replikert samtidig slik at det lages mange bakterier som inneholder profag. Bakterier med profag kalles lysogene celler. Plutselig når det blir gitt et signal kan fag lamda genomet gå ut av bakteriegenomet og starte en lytisk syklus. Sirkulært lambda-DNA replikeres uavhengig av vertscellens DNA. Forskjellige typer stress f.eks. store temperaturendringer, UV-lys etc. gjør at den lysogene fasen blir lytisk. Bakterier som gir difteri, skarlagensfeber og botulisme inneholder profag-gener som lager toksiner. Fra Corynebacterium er det kjent at en toksinproduserende stamme inneholder en temperat fag, men ikke-toksinproduserende stamme inneholder ikke noen bakteriofag. Clostridium botulinum med bakteriofag produserer toksin.
Bakteriofager kan brukes til å bekjempe bakterier, i stedet for å bruke antibiotika. Fykovirus er virus som infekterer alger. Som forsvar mot bakteriofagene har bakteriene utviklet restriksjonsenzymer som er utviklet ved naturlig seleksjon, slik at bakteriens eget DNA er beskyttet, mens fremmed DNA blir kuttet i biter av restriksjonssenzymene.
Historien om bakteriofager
Frederick William Twort (1877-1950) oppdaget bakteriofager i 1915, dvs. virus som kunne angripe og drepe bakterier. Dette ble uavhengig bekreftet av bakteriologen Felix d´Herelle (1873-1949) ved Pasteurinstituttet i 1917. Ved feltsykehus hadde d´Herelle sett soldater med dysenteri (blodgang), og d´Herelle dyrket opp bakteriene. Bekterieløsningen ble filtrert gjennom et bakterietett filter, og når filtratet ble blandet med nye bakterier ble bakteriene oppløst. Bakteriofagene formerer seg sammen med bakteriene. Selv om bakteriofagene er små, er de lette å påvise ved at de danner plakk på skåler med bakterier. Bakteriofagene er bakteriespisere. Bakteriofagterapi mot bakterieinfeksjoner er en metode som ble mye brukt i Øst-Europa, og har fått fornyet interesse grunnet antiobiotikaresistens.
Max Delbrück (1906-1981) som arbeidet som assisent hos Lise Meitner i Berlin, men måtte flyktet fra Nazityskland i 1937 til USA, Alfred Hershey og Salvador Luria mottok nobelprisen i fysiologi eller medisin i 1969 «for oppdagelser omkring replikasjonsmekanismen og genstrukturen hos virus».
Luria-Delbrück eksperimentet i 1943 med fag T1 som infekterer bakterien Eshcerichia coli viste at at bakteriers resistens mot virusinfeksjon skyldes tilfeldige mutasjoner og naturlig seleksjon ifølge Darwins evolusjonsteori. Genetiske mutasjoner i bakterier skjer i fravær av seleksjonspress, i dette tilfellet fag T1. Resistensen mot fag T1 eksisterte i populasjonen og ble ikke indusert ved eksponering for fag T1.
Bakteriofaggruppen Delbrück, Hershey og Luria hadde et undervisningstilbud i bakteriofaggenetikk ved Cold Spring Harbor Laboratory, New York i 1945, og ga et viktig grunnlag for utviklingen av molekylærbiologien hvor bruk av bakteriofager viste seg å være meget nyttig.
Bakteriefager er et viktig verktøy og gir økt forståele for molekylære mekanismer innen molekylærbiologi og genetikk.
Les mer i Historien om mikrobiologi og Historien om genetikk.
Peptidkommunikasjon mellom bakteriofager
Det er oppdaget at temperate fag, som veksler mellom forskjellige livssyklus: en lytisk fase eller en lysogen fase (latent hvilestadium), kan kommunisere med hverandre via peptider, et system kalt arbitrium. Peptider gir signal om at bakteriofagen skal gå over i lysogen syklus. I bakterier av slekten Bacillus har man identifisert genetisk mobile elementer med arbitrium. I tillegg til peptider kan arbitrium være quorum-signaler som N-acyl-homoserinlakton.
Litteratur:
Stokar-Avihail A,Tal N,Erez Z,Lopatina A & Sorek1 R: Widespread Utilization of Peptide
Communication in Phages Infecting Soil and Pathogenic Bacteria.. Cell Host & Microbe 25 (2019) 746-755. DOI:https://doi.org/10.10167/j.chom.2019.03.17
Hershey-Chase eksperimentet
De første studiene av bakteriofager ble gjort av den franske mikrobiologen Felix d'Hérelle (1873-1949) som arbeidet ved Pasteurinstituttet i Paris, og av den engelske bakteriologen Frederick Twort (1877-1950). Alfred Hershey og Martha Chase ved Cold Spring Harbor Laboratory, NY, i kunne i 1952 vise at det var DNA fra bakteriofagen T" som infekterte bakterien Escherichia coli og overtok bakteriens synteseapparat. Kappeproteinet i bakterien ble merket med radioaktivt svovel-35 (35S, med betastråling) som ble inkorporert i aminosyrene cystein og metionin i proteinet. DNA i bakteriefagen ble merket med fosfor-32 (32P). Det var P-32 merket DNA som ble funnet i bakterien, ikke S-35 merket protein.