Mykologi er det vitenskapelige fagområdet som omhandler sopp. Sopp er en heterogen gruppe organismer, fra encellete gjærsopp til flercellete, og spiller en fundamental rolle som i det biologiske kretsløp i alle økosystemer i vann og på land. Noen sopparter lever i mutualisme med alger og blågrønnbakterier (lav), noen i vomma på drøvtyggere, i fordøyelsesystemet til protister, eller i mutualisme inne i planterøtter (arbuskulær endomykorhiza) eller på utsiden av røttene på busker og trær(ektomykorrhiza ). Soppene er nedbrytere, noen lever som kommensaler, mens andre er patogener på dyr eller planter. Slekten Arthrobotrys er en predator som angriper nematoder i jorda.
Sopp er heterotrofe saprfofytter og absorberer næring fra omgivelsene. Mennesket bruker sopp og gjærsopp til en rekke nytteformål som mat, i næringsmiddelproduksjon og til å lage antibiotika og andre nyttige kjemiske stoffer. Blågrå østerssopp (Pleurotus ostreatus) er en østerssopp som dyrkes kommersielt på en blanding av halm, sagflis, og hestemøkk eller annet organisk materiale ved høy luftfuktighet e.g. inne i plastposer. Østerssoppen lever som saprofytt, men ikke i symbiose som mykorrhizasopp. Sjampinjonger (Agaricus) kan dyrkes til matbruk på samme måte. Til forskjell fra steinsopp og kantareller er avhengig av levende trær for å kunne vokse.
Sopphyfer
Sopphyfene er vanligvis delt i celler atskilt av tverrvegger (septa, entall septum). Sopphyfene danner filament- og rørformete celler omgitt av en plasmamembran, og en cellevegg med kitin. Et nettverk og samling av sopphyfer kalles et mycel. Imidlertid finnes det noen sopp som mangler tverrvegger (septa) kalt coenocytiske sopp med hvor en sopphyfe inneholder et stort antall cellekjerner, på lignende måte som hos amøber. Sopphyfene har cytoplasmastrømning og vokser i spissen av hyfene, og inntar ledige nisjer med organisk materiale. Når sopphyfene absorberer næring fra organiske stoffer i omgivelsene så senkes vannpotensialet i sopphyfene, vann følger etter og gir et turgortrykk. Celleveggen motstår turgortrykket og gjør at soppcellene ikke plasmolyserer, på lignende måte som hos planteceller. Mesteparten av det flercellete soppmycelet hos hattsoppene befinner seg som et stort nettverk nede i jorda. Mange av hattsoppene lever mutualistisk på røttene til trær hvor soppen mottar fotosynteseprodukter fra treet, mens soppen bidrar med opptak av mineralnæring til treet og beskytter planterøttene mot patogener.
Cellekjernene i sopphyfene er i de fleste stadiene haploide (n). Soppene formerer seg med sporer som enten kan være aseksuelle ved vegetativ formering eller kjønnet formering. Noen sopp slik som muggsopp kan bare formere seg aseksuelt, mens andre kan ha både vegetativ og seksuell formering. Ved seksuell formering blir det dannet seksuelle sporer fra fusjonering av cellekjerner fra to hyfer av kompatible krysningstyper. Cytoplasma i de to hyfene forenes med plasmogami, men de to cellekjernene fra de to krysningstypehyfene sameksisterer som et heterokaryon, altså et dikaryotisk mycel med to typer haploide cellekjerner. De to cellene vil etter en tid fusjonere ved karyogami og danne en diploid zygote. Zygoten er det eneste diploide stadiet hos sopp. Zygoten gjennomgår reduksjonsdeling (meiose) med rekombinasjon av gener som lager variasjon i de seksuelle sporene som evolusjonen kan selektere på. Soppsporene kan bli spredd over store avstander med vind eller vann, eller de kan bli fraktet med insekter og andre dyr.
Sopp og evolusjon
Sopp har flere utviklingsrekker gjennom evolusjonen. De monofyletiske gruppene sekksporesopp (askomyceter), stilksporesopp (basidiomyceter) og arbuskulære sopp (glomeromyceter), samt de polyfyletiske gruppene chytridesopp og koblingssopp (zygomyceter). Slekten Rozella er en gruppe chytridesopp som har atskilt seg fra andre chytridesopp tidlig i evolusjonen, og kanskje hører hjemme i en egen klad kalt Cryptomycota. Man antar at sopp gjennom evolusjonen har utviklet seg fra encellete protister i vann, utstyrt med flageller, en type amøber, nuklearider, som spiste alger og bakterier. Nålevende sopp har ikke flageller, bortsett fra hos en gruppe primitive chytridesopp. Flercellete dyr utviklet seg fra choanoflagellater, også en type protister, men i en uavhengig utviklingslinje forskjellig fra sopp, med atskillelse mellom sopp og dyr for ca. en milliard år siden. Man antar at de første terrestre sopp som dannet et grønt slim sammen med blågrønnbakterier etablerte seg på land før plantene. Men planter og sopp har en felles over 400 millioner år lang evolusjonshistorie.
Soppene er ordnet systematisk etter hvilken form for kjønnet formering de har. Asci hos sekksporesoppene (ascomycetene), basidier hos stilksporesopp (basidiemycetene), koblingssporerer (zygosporer) hos koblingssoppene, og meiosporer med en bakoverrettet flagell hos algesoppene (oomycetene).
Eldre systematikk baserer seg på hvordan fruktlegemene er utformet. De to viktigste avdelingene eller rekkene av soppriket er Ascomycota (sekksporesopp) og Basidiomycota (stilksporesopp). Myxomyceter (slimsopp) og plasmodiophoromyceter, som tidligere var systematisk plassert i soppriket, hører nå med til riket Protozoa. Disse har i stedet for mycel en naken amøbeformet flerkjernet kropp kalt plasmodium.
De organismene som tidligere hørte hjemme i gruppen algesopp (Phycomycota) er nå fordelt fire avdelinger eller rekker:
1) Rekke Oomycota (eggsporesopp), nå plassert i Protoctista eller Chromista som Pseudofungi).
2) Rekke Chytridiomycota (chytridesopp, algesopp, flagellsopp), eneste sopp med flageller festet ved basis til kinetosomer).
3) Rekke Hyphochytridiomycota (plassert i Protoctista eller i Chromista som Pseudofungi)
4) Rekke Zygomycota (koplingssopp). Parasittiske eller saprotrofe sopp. F.eks. slekten krypemuggsopp (Rhizopus).
5) Rekke Glomeromycota som omfatter endomykorrhizasopp.

Det finnes mer enn 1.6 millioner sopparter, mange lever i spesielle økologiske nisjer, og mange arter er kryptiske og uoppdaget. Sopp formerer seg med sporer som består av en eller noen få celler. Sporene kan formere seg ved deling eller ved knoppskyting som hos gjærsopp. Hos noen sopp lages aseksuelle sporer i et sekkformet sporangium og hos noen sopp kan sporene bevege seg med flageller (zoosporer), ofte hos de mer primitive. Andre sopp lager aseksuelle sporer kalt konidiesporer (konidier, gr. konis - støv) ved å avsnøre og kutte av ende- eller lateralceller fra spesielle hyfer kalt konidioforer. Aseksuelle konidiesporer kan lages enten nakent på konidioforene eller på innsiden av tykkveggete strukturer kalt pyknidier (gr. pyknos - tett). Hos noen sopp kan fragmenter av mycelet i form av terminale eller interkalære celler i hyfene forstørres og omgis av tykk vegg og kalles chlamydosporer (gr. chlamys - kappe; sporos - frø). Seksuell formering kan skje ved at to celler (gameter) med lik størrelse og utseende forenes og lager en zygote kalt zygospore. I andre sopp vil bevegelige gameter med lik eller ulik størrelse fusjonere og danne meiosporangier. Hos andre sopp er det ikke bestemte gameter som fusjonerer, men selve mycelet.
Sekksporesopp (Ascomyceter)
Hos sekksporesoppene (ascomycetene, Ascomycota) lages vanlig åtte seksuelle sporer i en sekklignende zygotecelle kalt ascus (gr. askos - sekk, pose) og sporene kalles ascosporer. Ascus (f.t.asci) eller det seksuelle stadiet kalles teleomorf (gr. teleos - fullstendig; morphe - form) eller perfekt stadium, mens konidie eller det aseksuelle stadiet kalles anamorf (gr. ana - bakvendt, baklengs) eller imperfekt stadium. Det perfekte stadiet er ofte det som overvintrer, men soppen kan også overvintre som mycel eller konidier. Ascus dannes etter befruktning av en hunnlig kjønnscelle (ascogonium) med en hannlig kjønnsspore antheridium (spermatium). Det befruktede ascogoniet lager ascogene hyfer som inneholder en hannlig og en hunnlig kjerne. Cellen på spissen gir en ascus. To kjerner fusjonerer og lager en zygote. Etter meiose dannes 4 haploide kjerner som etter en mitose gir 8 haploide kjerner. Disse omgis av vegg og danner ascosporer. Asci kan lages enkeltvis eller i grupper i form av et begerformet fruktlegeme kalt ascokarp. Mellom asci i et hymenium står sterile parafyser. Ascokarpen kan være en helt lukket beholder, kleistothecium (gr. kleistos - lukket; theke - boks), som hos f.eks. melduggsopp . Den kan være mer eller mindre lukket med en åpning (perithecium, gr. peri - rund) eller som en åpen flate i skåler eller beger (apothecium, gr. apo - bort fra). Ascomycetene identifiseres etter hvordan ascokarpen ser ut. Slekten penselmugg (Penicillium) infekterer matvarer, og sporene kan gi allergi. Roquefortmugg (Penicillium roqueforti) en en saprotrof sopp brukt til å lage blåmuggost og bidrar til blågrønne fargeflekker og streker, samt lukt. Noen har fått navn etter ostetypen som P. stilton og P. gorgonzolae.

Spissmorkel (Morchella conica) er en sekksporesopp som kommer med fruktlegeme tidlig om våren. Foldet apothecium. Innholder giftstoffet gyromitrin.
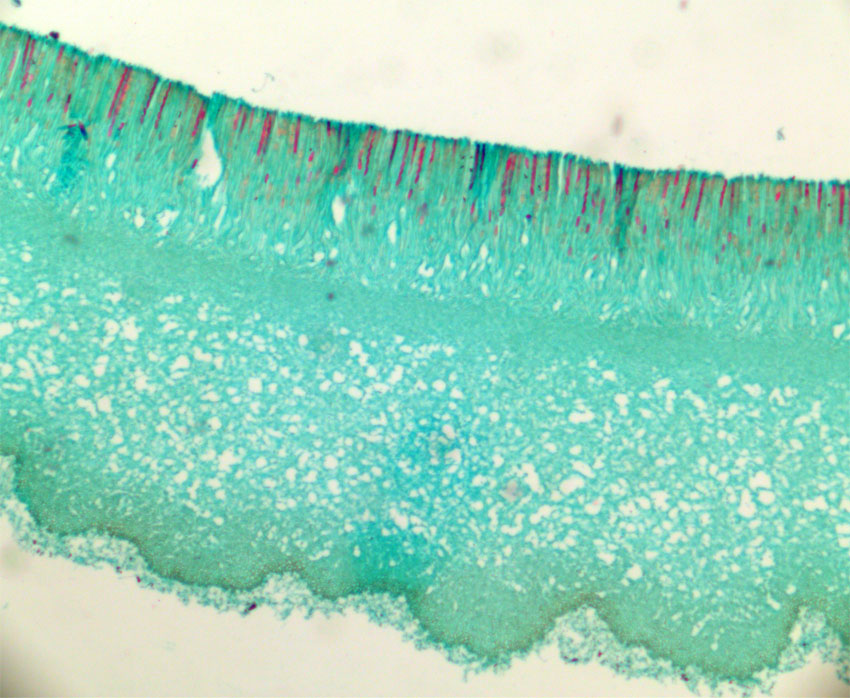
Bilde. Tverrsnitt av skarlagenvårbeger (Sarcoscypha austriaca). Sekksporer (asci) på oversiden.
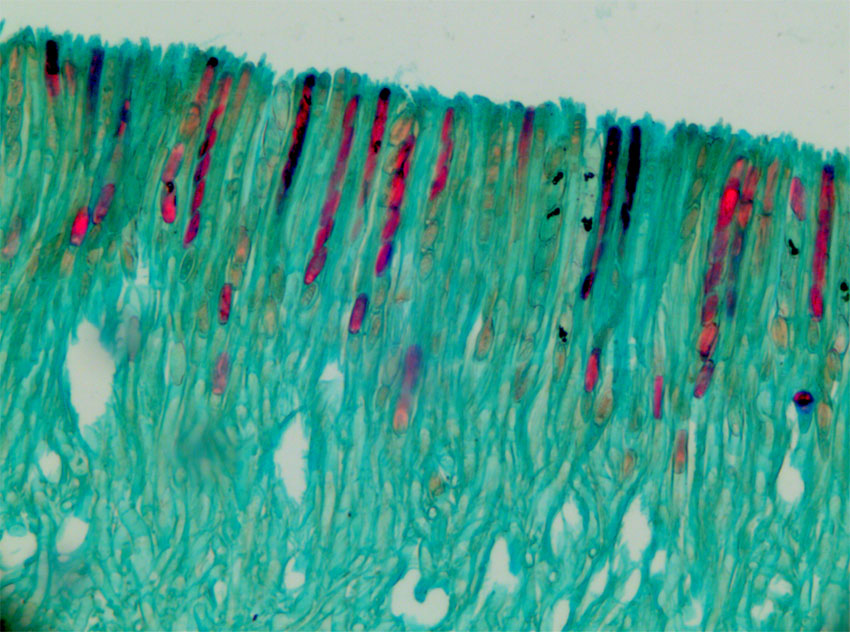
Bilde. Tverrsnitt av skarlagenvårbeger (Sarcoscypha austriaca). Detalj fra sekksporer (asci). Åtte ascosporer i en ascus.

Lønntjæreflekk (Rhytisma acerinum) på spisslønn er en biotrof ascomycet i orden Rhytismatales gir utover sommeren tjærefargete flekker (teleomorf-stadiet) omgitt av en gul rand med klorose. Om våren fra overvintrede blad med apothecier lages nåleformete ascosporer som spres med vinden. Sporer som lander på spisslønnblad spirer og hyfen vokser gjennom spalteåpningene og inn i bladet, og hvor man kan observere gulaktige flekker. På ettersommeren i de svarte flekkene finnes anamorf-stadiet Malasmia acerina med acervulus (fl.t. acervuli) lager konidioforer som lager konidier
Stilksporesopp (Basidiomyceter)
Hos stilksporesopp (Basidiomycetes gr. basidie - grunnlag, sokkel; mykes - sopp, Basidiomycota) lages seksuelle basidiosporer på utsiden av en klubbeformet zygotecelle kalt basidium. Basidiosporene kan sitte på sterigmer (stilker). Fruktlegemet (basidiokarpen) er ofte utformet som en hatt eller trakt, med basidiene (sporangiene) på undersiden. Hattsoppene er basidiomyceter og hatten lager på undersiden et hymenium (gr. hymen - skinn, hud, hinne) hvor endecellene er basidier med basidiesporer. Mellom basidene kan det være beskyttende sterile cystider.
Hymeniet er utviklet som skiver eller ribber (skivesopp), rør (rørsopp), pigger (piggsopp), porer (poresopp) eller greiner (korallsopp og fingersopp) eller kan være glatt (barksopp).

Gullgaffel (Calocera viscosa), Basidiomycota orden Dacrymycetales er en gelesopp med gelatinøst og slimete utseende.

Gul gelesopp (Tremella mesenterica) i orden Tremellales vokser på døde kvister og greiner fra løvtrær. Gelatinøst guloransje fruktlegeme som kommer fram i fuktig vær (heksesmør).
Røyksopp mangler hymenium, men har basidiene inne i fruktlegeme i en gleba (sporemasse). Hymeniet er noen ganger dekket av et slør (hylster, velum partiale). Basidiesopp kan danne ektomykorrhiza, men det fleste er saprotrofe, noen lever parasittisk og noen danner lav sammen med grønnalger.

Stilksporesoppen og Bbasidiomyceten Exobasidium vaccinii på tyttebær (Vaccinium vitis-Idaea) . En parasittsopp (Exobasidiomycetales) på arter i lyngfamilien (Ericaceae). Soppen lager deformerte blad med rose eller purpurfarget overside, og hymeniet er hvitfarget filtaktig på undersiden av bladet.
Rustsopp
Rustsopp, som også hører med til Basidiomycetene, har ikke fruktlegemer. Istedet kan rustsopper ha fem forskjellige fruktstrukturer og fem forskjellige sporetyper. Noen av sporene kan infektere bare en vert. Alle rustsopp lager teliosporer (teleutosporer, gr. teleute - fullføring, avslutning) og basidiosporer. I tillegg kan det produserers spermatier (pykniosporer), aecidiosporer (l. aecidium - beger, kopp, vårsporer) og uredosporer (l. uredo - rust, meldugg). Teliosporerene fungerer som seksuelt overvintringsstadium som ved spiriring om våren lager et basidium. Spermatier er hannlige gameter, men kan ikke infektere planter. De fertiliserer mottakerhyfer med rett krysningstype (+,-) og lager et dikaryotisk mycel med dikaryotiske sporer. Dette mycelet lager aeciosporer, som etter infeksjoner lager mer dikaryotisk mycel som nå lager uredosporer (sommersporer). Et sklerotium er tettpakket hyfevev med en hard ytre vegg og fungerer som overvintringsorgan.
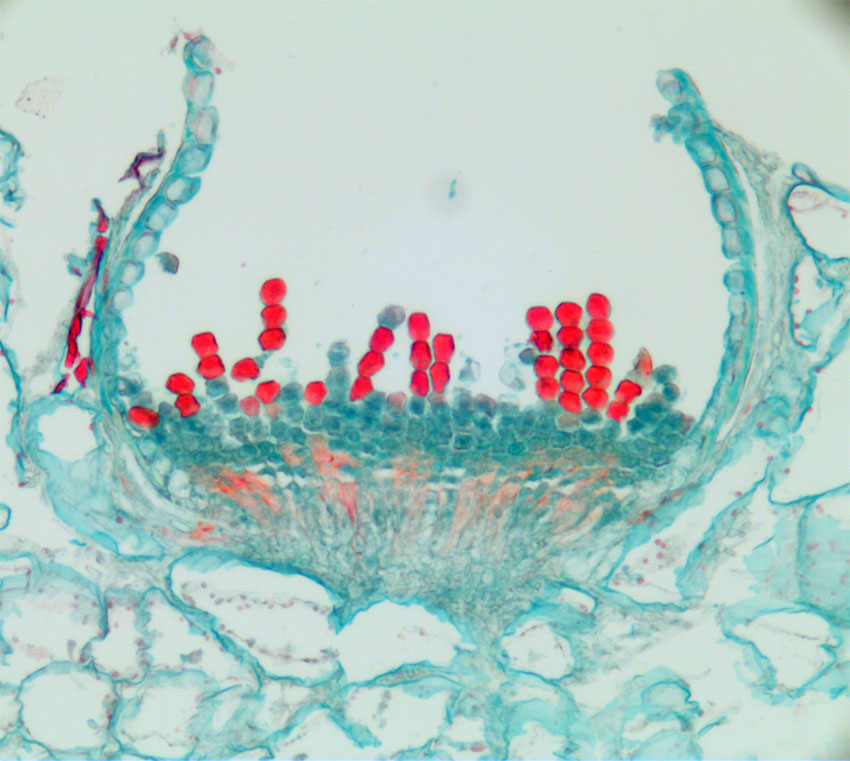
Bilde. Rustsoppen Puccinia poarum har aecidier med aecidisporer på undersiden av blad hos hestehov (Tussilago farfara), sees som oransjefargete flekker. Rustsoppene er parasitter på planter, og mange av dem har vertsskifte. Bilde viser et spermogonium, skålrust med rekker av aecidisporer og på hver side en rekke med tykkveggete peridieceller . Sporene blir spredd med vinden.
Svartrust
Svartrust Puccinia graminis er en biotrof rustsopp som har alle 5 sporestadiene, og vertsveksler (heteroik) med berberis (Berberis vulgaris). Svartrust angriper flere av kornslagene. Siste stadiet om høsten utvikler tykkveggete dikaryote svartfargete vintersporer (teliosporer), derav navnet. Svartrust har forskjellige patovarer (pv.). Puccinia graminis f. sp. tritici angriper hvete (f. sp. = forma specialis). P. graminis f. sp. hordei angriper bygg, og P.graminis f. sp. avenae angriper havre osv. I foredlingsarbeid utvikles sorter som er resistente mot svartrust, og man kan sprøyte med fungicider, men Ug99 (TTKSK), som ble oppdaget i Uganda i 1999, omgår Sr resistensgenene og truer nå hveteavlingene i Afrika. Om sommeren produserer soppmycelet på hveteplantene dikaryotiske (to haploide kjerner i en celle) sommersporer (uredosporer, uredionosporer) fra uredinier. Uredionosporer infekterer andre hveteplanter, og kan bli spredd over store områder. Dikaryote teliosporer infekterer berberis. Det skjer karyogami hvor kjernene fusjonerer, og ved reduksjonsdeling (meiose) dannes fire haploide basidiosporer fra hver teliospore. På bladene til berberis spirer basiodiosporene, trenger gjennom epidermis og det haploide mycelet inntar bladet. Dette mycelet lager en struktur kalt pyknier, hvor det blir dannet haploide sporer kalt pyknosporer og reseptive hyfer. Pyknosporene er omgitt av et klebrig honningdugg til tilrekker insekter som sprer sporene fra blad til blad, men sporene kan også bli spredd med plaskregn. Pyknosporene kan fusjonere med reseptive hyfer av rett krysningstype og danne et dikaryot mycel. Det er i dette seksuelle stadiet soppen har genetisk rekombinasjon og hvor det kan utvikles nye patovarer av soppen. Det dikaryote mycelet danner aecier som lager kjeder med vårsporer, aeciosporer omgitt av sopphyfer. Aeciosporene blir overført til hveteplanter eller andre kornslag, hvor sporene spirer og infekterer. Det diakaryote mycelet produserer etter hvert uredosporer og ringen er sluttet. Allerede i 1916 ble det i Norge forbudt å dyrke Berberis vulgaris, og de andre sortene berberis som nå brukes i hager er resistente mot soppen.
Filtrust
Filtrust (Cronartium ribicola) kan observeres som oransje sommersporer under solbærbladene, og bladene felles tidligere enn vanlig om høsten. Filtrust kan også angripe andre bærsorter i slekten Ribes. Om høsten dannes vintersporer som gir brunfargete tråder med basidiesporer som overfører til femnålete furuarter som soppen vertsveksler med. På furu bryter det ut aecidiesporer gjennom barken.

Filtrust (Cronartium ribicola) på undersiden av solbærblad sorten Øyebyn. I dette stadiet spres sporene med vinden over til andre blad-

Filtrust (Cronartium ribicola) sett fra oversiden av solbærblad sorten Øyebyn.
Eggsporesopp (Oomyceter)
Sopplignende oomyceter har gametangier av ulik størrelse som fusjonerer til en zygote kalt oospore. Livssyklus hos sopp: Sporestadiet er vanligvis haploid (n). Sporen spirer til en hyfe med haploide kjerner. En hyfe kan lage haploide sporer igjen, som hos imperfekt sopp, eller smelte sammen med en annen hyfe hvor kjernene fusjonerer og danner en diploid kjerne kalt zygote. I de fleste stilk- og sekksporesopp vil de to kjernene i den fertiliserte hyfen ikke fusjonere, men være atskilt i cellen i par, og som deler seg og danner hyfer med to kjerner. Hos sekksporesoppene vil de dikaryote hyfene finnes bare på innsiden av fruktlegemet hvor de blir ascogene hyfer hvor to kjerner fusjoneres til zygote som deler seg meiotisk og produserer ascosporer med haploide kjerner. Hos stilksporesoppene lages haploide sporer bare på korte haploide hyfer. Dikaryot vev (n + n) gir fruktlegemet. Dikaryote hyfer kan produsere aseksuelt dikaryot mycel. Når det dannes en zygote kan det ved meiose lages basidiesporer. Sopp lager aminosyren lysin via mellomproduktet alfa-aminoadipinsyre.
Den vanligste soppen som brukes til menneskemat i vestlige land er sjampinjong (Agaricus bisporus), og i østen shiitake (Lentinus edulus).
Sekundærmetabolitter hos sopp
Sekundærmetabolitter er kjemiske stoffer laget i biosynteseveier, og som ikke er en del av primærmetabolismen. Flere av sekundærmetabolittene virker som gift (mykotoksiner) på virveldyr (soppgifter). Polymerer av acetat i form av acetyl-CoA gir opphav til polyketider, bisfuranokoumariner og spesielle fettsyrer. De lange kjedene kan endres ved ringdannelse, eller ved å danne tioestere, amider eller laktoner.
Aflatoksiner er polyketider laget av Aspergillus-arter. Aflatoksin B1 er meget giftig (aflatoksikose), også karsinogen, og blir laget i soppinfisert korn og nøtter oppbevart fuktig ved høy temperatur. Husdyr som høns, kalkuner, ender, griser og kveg som spiser Aspergillus-infisert for, kan føre giften videre i næringskjedene, e.g. i form av aflatoksin M.
Zearalenon er en syklisk polyketode laget i shikimat- eller polyketide-biosynteseveien, blant annet laget av Fusarium graminearum og andre Fusarium-arter. Zearalenon virker som en hormonhermer. Zearalenon sammen med andre mykotoksiner som T-2-toksin, HT-2-toksin, sporodemin, og vomitoksin (deoxynivalenol, et trichothecen og epoksysesquiterpen) kan gi forgiftninger via korn blant annet brukt i kyllingoppdrett.
Patulin er en polyketide laget i acetat-malonat biosynteseveien, og finnes blant annet i soppinfiserte epler, produsert av Penicillium expansum, katalysert av et stort enzymkompleks Isoepoksydon dehydrogenase Et av mellomproduktene i biosynteseveien er 6-metyl-salicylsyre.
Amatoksiner er sykliske peptider som påvirker syntesen av RNA hos eukaryoter og finnes i fluesopp (Amanita).
Mykotoksiner som blir laget fra aminosyrer er bufotenin fra fluesopp (Amanita) og psilocybin fra felinsopp (Psilocybe).
Noen sopp lager sekundærmetabolitter som virker antibiotika. Ergot-alkaloider blir laget av meldrøyesopp fra aminosyren tryptofan i shikimatbiosyntesevien sammen med isoprenoider fra mevalonatbiosyntesevien. Soppen Gibberella lager gibberelliner som virker som plantehormon.
Noen bioaktive sekundærmetabolitter ekstrahert fra piggsvinsopp (Hericium erinaceus), i klassen Agaricomycetes i Basidiomycota har mulige positive helseeffekter på flere nevrodegenerative sykdommer Alzheimers sykdom, Parkinsons sykdom og depresjon, men man må ikke la seg lure. Eksempler på slike stoffer er erinaciner type A-K, P og Q),en type diterpenoider og hericenoner C-H som er benzaldehyder.
Piggsvinsoopp (Lions mane) (Hericium erinaceus, l. erinaceus – piggsvin) orden Russulales, familie Hericiaceae.anvendt i Øst-Asia og inngår i kinesisk medisin
Historie
En italienske botanikeren Pier Antonio Micheli (1679-1737) oppdaget sporer hos sopp og hadde gode kunnskaper om kryptogamer. Han viste at sopp ikke kan oppstå spontant, men spres med sporer. I Nova plantarum genera iuxta Tournefortii methodum disposita (1729) ga han beskrivelser av mange planter, sopp og lav. Micheli var professor ved Universitetet i Pisa og leder av den botaniske hagen i Firenze.
Den svenske botanikeren og mykologen Elias Magnus Fries (1794-1878) med verkene Observationes mycologici og Systema mycologicum i tre deler (1821-1832) innehold grunnleggende informasjon om sopp på den tiden, autornavn Fr. Han introduserte sopp som mat med Anteckningar öfver de i Sverige växande ätliga svampar i 1836 og plansjeverket Sveriges ätliga och giftiga svampar. Han samlet også sopp til fytoteket i Uppsala. Fries organiserte sopp i den eldre taksonomiske gruppen hymenoceter (Hymenocetes) som bl.a. omfattet ordnene skivesopp (Agaricales), rørsopp (Boletales) og kremler (Russulales).
Den britiske mykologen Cecil Terence Ingold (1905-2010) ga navn til den eldre betegnelsen ingoldske sopp, akvatiske sopp med stjerneformete sporer innen orden Pleosporales, klasse Dothideomycetes, som infekterer fisk. Disse soppene har etter DNA-sekvensering nå fått nye navn.
Den franske legen og mykologen Charles Tulasne (1816-1884), med autornav C.Tul., utga den illustrerte Selecta Fungorum Carpologia og broren den franske botaniker og mykologen Louis René Tulasne (1815-1885) bidro blant annet til beskrivelse av hypogeiske ascomycetsopp med underjordiske fruktlegemer og trøffelstrategi plassert i orden Tuberales. Disse har evolusjonære likhetstrekk med de koppformete fruktlegemene man finner i orden begersopp (Pezizales) og seinere blitt flyttet til denne orden med familiene Discinaceae, Glaziellaceae, Helvellaceae, Pezizaceae, Pyronemataceae og Tuberaceae, hvorav bare den siste familien med hypogeiske fruktlegemer som ikke har eksplosiv mekanisme for sporespredning. Trøffelslekten Tuber har fruktlegemer som minner om poteter. Slekten Geopora i Pyronemataceae har vekststrategi som minner om trøfler, men på slutten av utviklingen bryter fruktlegmet gjennom jordoverflaten har aktiv sporespredning over bakkenivå.
Mer om sopp og gjær
Litteratur
Wikipedia