Antioksidantforsvar
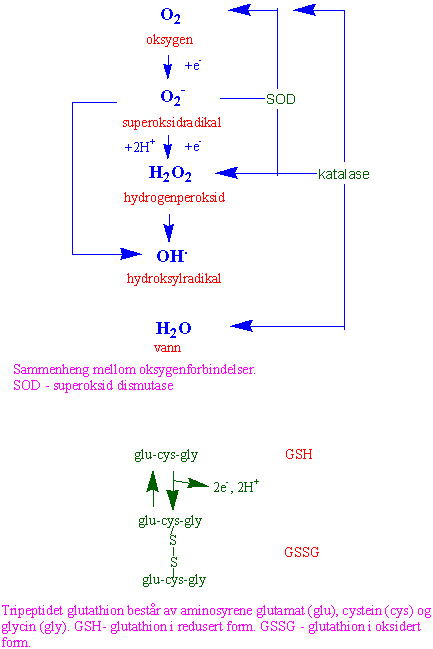
For å beskytte seg mot de skadelige effektene av ROF har alle organismer et aktivt antioksidantforsvar bestående av vitaminer (vannløselig vitamin C (askorbinsyre) og fettløselig vitamin E (alfa-tokoferol), tripeptidet glutation (GSH), enzymer (superoksid dismutase, katalase, askorbat peroksidase, glutation peroksidase, glutation reduktase og peroksiredoksin), bl.a. i askorbat-glutation syklus, karotenoider, samt anthocyaniner ROF kan også brukes aktivt i forsvar mot andre skadelige og sykdomsfremkallende organismer som virus, bakterier, sopp og beitende dyr, bl.a. katalysert av NADPH oksidase:
NADPH + 2O2 ⇒ NADP+ + 2O2·- + H+
Reaktive oksygenforbindelser kan også delta i ordinære signalveier i metabolismen. Hydrogenperoksid og NO deltar i en rekke signalveier i plantene som aldring, lukking av spalteåpninger, frøspiring,blomstring og rotdannelse. Hydrogenperoksid kan bli laget både enzymatisk og ikke-enzymatisk.
Hydrogenperoksid kan i reaksjon med fritt jern gi Fenton- og Haber-Weiss-reaksjoner som danner det meget reaktive og skadelig hydroksylradikalet (OH·). For å hindre slike Fenton-reaksjoner er jern alltid hard bundet enten i hem eller i jern-svovelforbindelser, eller jern er chelatert til askorbinsyre under transport.
Anthocyaniner er en gruppe flavonoider som i tillegg har funksjon som beskyttelse mot UV-stråling, signalmolekyl som lokker til seg pollinatorer, frukt- og frøspisere, men blir også dannet under biotisk stress som næringsmangel, spesielt ved mangel på nitrogen og fosfor, ved saltstress og ved høy lysintensitet. Ved slikt stress dannes det reaktive oksygenforbindelser og man tenker seg at anthocyaniner er en del av antistressforsvaret.
I vann-vann-syklus i fotosyntesen (Mehler-reaksjon) blir det dannet superoksidanionradikaler når det elektronegative oksygen tiltrekker seg elektroner som kommer fra vann i tilknytning til det oksygenutviklende komplekset i fotosystem II. Superoksidantionradikalet blir fjernet via enzymene superoksid dismutase og askorbat peroksidase, og det blir dannet vann.
Superoksidanionradikalet kan også bli dannet mellom kompleks I og III i elektrontransportkjeden i mitokondriene.
I fotorespirasjon når glykolat omdannes til glyoksylat katalysert av glykolat oksidase, og ved nedbrytning av fettsyrer via beta-oksidasjon blir det dannet hydrogenperoksid. Disse reaksjonene er derfor lokalisert til peroksysomer og glyoksysomer og hydrogen peroksid blir fjernet via enzymet katalase.
Ved polymerisering og dannelse av lignin i celleveggene deltar hydrogenperoksid og celleveggperoksidase.
Hydrogenperoksid (H2O2) deltar sammen med plantehormonet abscisinsyre i signalveien som fører til lukkingen av spalteåpninger på blader ved tørke. Det samme gjelder en signalvei via hydrogenperoksid, heterotrimere G-proteiner og plantehormonet etylen som også fører til lukking av spalteåpningene.
Nitrogenoksid (NO) er et radikal som har en viktig signalfunksjon i dyr og planter. Både hydrogenperoksid og NO kan bli dannet under stress, og kan i samvirke med kalsium (Ca2+) gi bl.a. programmert kontrollert celledød og aldring.
Ozon blir dannes i stratosfæren, men også som bakkenært ozon i troposfæren i reaksjon med nitrogenoksider og flyktige organiske molekyler, inkludert luftforurensninger. Årsaken til at bakkenært ozon gir lukking av spalteåpninger på bladene er at ozon gir opphav til hydrogenperoksid i spalteåpningsapparatet.
Glukose oksidase (notatin) er et enzym som finnes bl.a. hos sopp og som oksiderer glukose til hydrogenperoksid og D-glukono-1,5-lakton:
\(\alpha-\)D-glukose ⇔ \(\beta-\)D-glukose + O2 ⇒ D-glukono-1,5-lakton + H2O2
Oksalat oksidase katalyserer reaksjonen hvor oksalsyre blir oksidert til karbondioksid og hydrogenperoksid:
oksalat + O2 + 2H+ ⇔ 2CO2 + H2O2
Sulfitt oksidase i peroksysomene i planter oksiderer sulfitt, bruker oksygen som elektronakseptor og det dannes sulfat og hydrogenperoksid. Sulfitt oksidase deltar i beskyttelse mot forurensning av svoveldioksid (SO2). Sulfitt oksidase finnes hos bakterier, planter, dyr og mennesker. Hos mennesker deltar sulfitt oksidase i oksidasjon og nedbrytning av svovelaminosyrene cystein og metionin, hvor sulfitt blir oksidert til sulfat. Sulfitt oksidase har et domene for binding av hem og et domene som binder molybdopterin.
{kind=link}