Røtter dannes fra det rotapikale meristemet i rotspisser og har til oppgave:
1) Feste planten til jorda.
2) Ta opp vann og næringssalter fra jorda.
3) Pppbevare opplagsnæring hos flerårige planter i form av rotknoller eller oppsvulmete hovedrøtter, siderøtter eller adventivrøtter.
Rota utvikler seg fra et frørot (kimrot) når frøet spirer, og kimrota er det første som trenger igjennom frøskallet. Hos mange arter slik som gras dør kimrota og blir erstattet av adventivrøtter (birøtter, tilleggsrøtter) som vokser ut fra den nedre del av stengelen og danner en knipperot (trevlerot). Hos andre utvikles kimrota til en hovedrot, ofte pelerot, og fra denne vokser det ut siderøtter (rotgreiner), som igjen kan gi opphav til nye siderøtter. Ved rotspissene er det en rothårssone med rothår.
Rota er fra spissen og oppover delt i følgende soner med flytende overgang mellom dem: delingssone, streknings- og differensieringssone og rothårssone. I delingssonen er det en stille sone hvor mitose skjer saktere enn i cellene omkring. Det apikale meristemet er beskyttet av en rothette. Rothettecelle løsner lett og skiller ut karbohydratslim som smører rota på sin vei gjennom jorda. Noen av cellene i rothetta, i kolumella, inneholder statolitter som responderer på tyngdekraften (gravitropisme).
Rothår (sugehår) er utposninger fra epidermisceller like bak delingssonen (meristemet). Rothårene tar opp vann og grunnstoffer i ioneform fra jorda. Rothårene danner store overflater og er svært viktige for plantenes vann- og næringsopptak. Vann med næringssalter følger apoplast eller symplast inn til endodermis.
Siderøtter dannes fra pericykel, et stykke lenger bak rotspissen. Rota består av epidermis, rotbark (cortex) og sentralsylinder med ledningsvev (xylem og floem) omgitt av pericykel og en endodermis med Casparysk bånd (suberin). Enfrøbladete røtter har en marg i sentrum av rota. Røtter hos tofrøbladete planter og gymnospermer har sekundær tykkelsesvekst via vaskulært kambium som danner sekundært vedvev (xylem) og silvev (floem), samt et korkkambium som danner sekundært hudvev (korkhud, periderm).
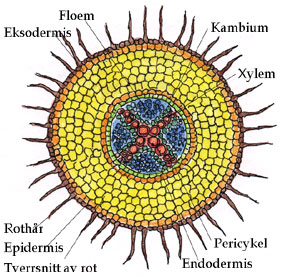
Tverrsnitt tofrøbladet rot. I en enfrøbladet rot er det en marg i sentrum av rota.
Den unge spirende frøplanten har en kimrot, og denne kan vokse og videreutvikle seg til en hovedrot (pelerot) hos de fleste tofrøbladete planter og gymnospermer. Hos de enfrøbladete erstattes kimroten av en rekke adventivrøtter (birøtter) som lages på stengelens nedre del og utvikler seg til en knipperot (trevlerot). Birøtter (adventivrøtter) utvikles generelt fra stengel eller blad, ikke fra en rot.
Kontraktile røtter (l. trahere - trekke) har ofte et rynket utseende og kan ved rotforkortning være med å trekke planten nedover i jorda og bringe overvintringsknopper ned til jordoverflaten eller under denne. Knoller, løk, og rhizomer som vokser vannrett kan også bringes til rett dybde i jorda ved rotforkortning. Det kan dannes rotknopper (biknopper) på røttene fra pericykel f.eks. hos småsyre og lintorskemunn. Hvis roten svulmer sterkt opp med mye opplagsnæring og liten forgreining kalles røttene ammerøtter. Det er en flytende overgang fra ammerøtter til det som kalles rotknoller hvor omtrent hele roten er svulmet opp med opplagsnæring.
Planter kan ha klatrerøtter (bergflette (Hedera helix)) eller klamrerøtter (Ficus-arter, trekveler (Clusia rosea)). I tropene finnes brettrøtter eller platelignende røtter ved basis av tree.
Støtterøtter kan vokse fra stamme eller greiner ned til jorda og har støttefunksjon (f.eks. mangrove (Rhizophora), banyantre (Ficus benghalensis)).
Ånderøtter sørger for oksygentilgang til røttene, vokser negativt gravitropisk og er vanlig i mangroveskog (Avicennia).
Daukiforme siderøtter (navn fra Daucus – gulrot) er korte (1-2 cm) oppsvulmete gulrotformete siderøtter med svært lange rothår, og rothårene kommer fra alle epidermiscellene. Finnes hos arter i halvgrasfamilien (Cyperaceae) som vokser på våtmark med lavt fosforinnhold.
Epifyttiske orkidéer har luftrøtter med en epidermis bestående av flere lag som kan suge vann kalt velamen (l. velamen - dekke, overtrekk).
Snyltende planter har snylterøtter med haustorier som trenger inn i vertsplanten. Alle røttene på en plante kalles rotsystemet. Ledningsvev i sentralsylinder (stele) ligger radiært i sentrum av roten. Avhengig av hvordan den første kimroten utvikles fra embryo kan plantene deles inn i allorhizofytter som omfatter gymnospermene og angiospermene hvor rotspiss og skuddspiss ligger motsatt av hverandre i hver sin ende av embryoaksen; og homorhizofytter som omfatter de laverestående plantene hvor den første rota anlegges lateralt i forhold til embryoaksen.
Greining av røttene skjer i god avstand fra rotspissen, hvor siderøtter dannes i modne områder som har sluttet å strekke seg. Røttene lages fra en rotmeristem. Apikalt rotmeristem er dekket av en rothette som lages av rothettestamceller. Rothetta beskytter meristemet på veien ned i jorda, slites av, og er ikke en permanent del av rota. Rothetta har en sentral sone kalt kolumella som inneholder statolitter som registrerer retningen for tyngdekraften. Et rolig senter er en sentralt plassert del av meristemet hvor cellene deler seg med en mye lenger tidssyklus. Hvis f.eks. cellesyklus varer i 400 timer i rolig senter, mens celledelingen (doblingstiden) tar bare 10 timer i rothetta betyr dette at en rothettecelle har delt seg 40 ganger og produsert 1.1 x 1012 celler, mens en celle i rolig senter bare er ferdig med en celledeling. Med radioaktiv tritium-merket thymidin vil celler som har DNA replikasjon og aktiv deling inkorporere radioaktiv thymin i kjernen. Ved mikroautoradiografi vil filmen bli svertet der hvor det er aktiv celledeling, mens den er blank i rolig senter.
Røttene er i spissen dekket av en rothette som beskytter det apikale rotmeristem. Rothetta skiller ut karbohydratslim (mucigel) som gir smøring slik at rota glir lettere gjennom jorda, beskytter mot uttørking, og letter overføring av mineraler. Organiske stoffer fra røttene som skilles ut i jorda gir en rik mikroflora og mikrofauna rundt røttene. og virker som et mikromiljø for mikroorganismer.
Det er overlappende soner: rothettesone, meristemsone med rotapikalt meristem som danner primærrot, strekningssone med raske cellestrekning, og noen celler fortsetter å dele seg, samt en modningssone med differensiering. Veksten er begrenset til rotspissen, inkludert spissen av siderøtter. I en sone bak rotspissen er endodermisceller utformet som rothår. Rothårsonen gir stort overflateareal på røttene.
Siderøtter (lateralrøtter) utvikles endogent fra pericykel innnenfor endodermis. Siderota vokser gjennom cortex og epidermis og lager en ny vekstakse
Når et frø spirer kommer rota først ut av frøet og danner en kimrot. Kimrota kan være varig som hos tofrøbladete planter, og gir basis for rotutviklingen og danner en pelerot med en hovedakse som vokser i tykkelse og fra denne vokser det ut siderøtter. Hos arter i soleiefamilien (Ranunculaceae) og hos enfrøbladete planter som f.eks. gras er kimrota lite varig og det utvikles en knipperot bestående av omtrent like store røtter med samme diameter som kommer ut fra basis av stengelen.
Røtter som utvikles fra en stengel kalles adventivrøtter. Navnet adventiv refererer seg til at de dannes på et uvanlig sted. Enfrøbladete planter har 3-6 primære rotakser som kommer ut fra frøet, og seinere dannes adventivføtter (nodale røtter) som greiner seg og gir knipperot. I røttene utvikles floem raskere enn xylem siden røttene har behov for assimilater fra fotosyntesen i skuddet for å kunne vokse.
Mangrovetrær bl.a. Avicennia og Sonneratia får tilført oksygen til røttene ved hjelp av ånderøtter (pneumatoforer) som vokser oppover mot og over vannoverflaten (negativ gravitropisk). Ånderøtter i hundrevis som snorkler i med porer og luftganger er forbundet med rotsystemet. Mangrove f.eks. Rhizophora har også støtterøtter. Mangrove i tidevannsbeltene langs kysten i subtropiske og tropiske områder hindrer erosjon, beskytter kysten og fanger opp leirsedimenter med ånde- og støtterøttene, og danner en viktig biotop for blant annet småfisk. Planter som lever i sjøvann opplever fysiologisk tørke. Disse må ha et lavere vannpotensial i rotcellene enn i vannet omkring røttene for å kunne ta opp vann. Arter som Avicennia og Aegiceras skiller ut overskudd av salt gjennom saltkjertler på bladene. Rhizophora, Lumnitzera og Sonneratia er ultrafiltrere som holder Na+ ute og har lite salt i xylemvæsken. Mangrovene må tåle endringer i saltinnholdet i vannet, bl.a. i regntiden med mye ferskvann fra slamrike elver. Mange mangrover kan imidlertid vokse i ferskvann bl.a. Bruguiera. Mangrovene Bruguiera og Ceriops har knerøtter som vokser horisontalt, men som lager med mellomrom en kneformet bøy over vannet. Disse virker både som ånde- og støtterøtter. Sumpsypress (Taxodium distichum) har også knerøtter. Mangrovetreet Xylocarpus granatum har plate- eller plankeformete røtter som vokser vertikalt.
Banyantreet (Ficus benghalensis), opprinnelig voksested i India og Pakistan, ofte plantet rundt Hindu- og Buddhist-templer, har luftrøtter som henger ned fra greinene. Disse blir røtter når de treffer bakken og virker som vertikale støtterøtter, noe som kan gi treet en betydelig omkrets. I tillegg har stammen brettrøtter. Støtterøtter laget fra luftrøtter finnes også hos gummitreet (Ficus elastica). Det fortelles at Aleksander den store hadde 7000 soldater i skyggen av ett Banyantre. Luftrøtter finnes også hos vindusblad (Monstera). Hos orkidéer som lever som epifytter i tropisk regnskog er luftrøttene dekket av en gråhvitfarget flerlaget epidermis med døde celler med fortykkete vegger, kalt velamen, som kan absorbere vann.
Kvelerfiken (Ficus aurea) og trekveler (Clusia rosea) har klebrige frø som spres med fugler. Frøene spirer høyt oppe i trærne i regnskogen og sender luftrøtter ned mot bakken. Det dannes en nettverk av røtter som kan vokse sammen og når disse når bakken vokser det opp en stamme på vanlig måte. Treet som kveleren klenget seg til blir skygget vekk og dør pga. rotkonkurranse.
Berglette (Hedera helix) og Philodendron har klatrerøtter på stengelen, altså adventivrøtter.
Hos mais virker adventivrøttene fra den nedre del av stengelen som støtterøtter.
Deler av rota kan utvide seg til en rotknoll med opplagsnæring. I noen tilfeller inngår deler av stengelen (hypokotyle) i rotknollen. Nytterotvekster er vanligvis pelerøtter med opplagsnæring fra toårige planter. Røtter som regulerer dybden den vokser med kan få en rynket overflate.
Noen planter er uten røtter som f.eks. spansk mose (Tillandsia usneoides) i ananasfamilien (Bromeliaceae). Navnet har den fått fordi den ligner på en lav (Usnea), og den henger ned fra greiner på samme måte. Den er grå i tørr tilstand og lys grønn når den er våt. Andre planter uten røtter er vannplanter som flyter i vannmassene f.eks. blærerot (Utricularia).
Rotoverflaten på en urtaktig plante er ca. 100-300 m2. En ørkenplante som Prosopis har røtter som går 50 meter ned til grunnvannet. Hos ettårige går rota fra 0.1-2 meter dypt og lateralt fra 0.3-1 meter. Evne til opptak og vann med mineralnæring er avhengig av et stort og velutviklet rotsystem.
Røtter kan være utformet som et oppsugingsorgan (haustorium) hos parasitterende planter f.eks. misteltein (Viscum album) og skjellrot (Lathraea squamaria).
ARABIDILLO er et gen hos Arabidopsis som koder for et F-boks protein som gir siderotdannelse og greining av røttene.
.jpg)
Bilde 1. Utsnitt av en rot av mais (Zea mays). Ytters epidermis bestående av ett cellelag. Under dette en hypodermis (eksodermis) som man antar har samme funksjon som endodermis. Deretter følger rotbarken (cortex) og endodermis med Casparysk bånd bestående av lignin. Endodermis består bare av ett cellelag. Innenfor endodermis ligger pericykel, bare ett cellelag, og innen for dette ligger ledningsvevet: xylem med tykke cellevegger og floem med tynne cellevegger.
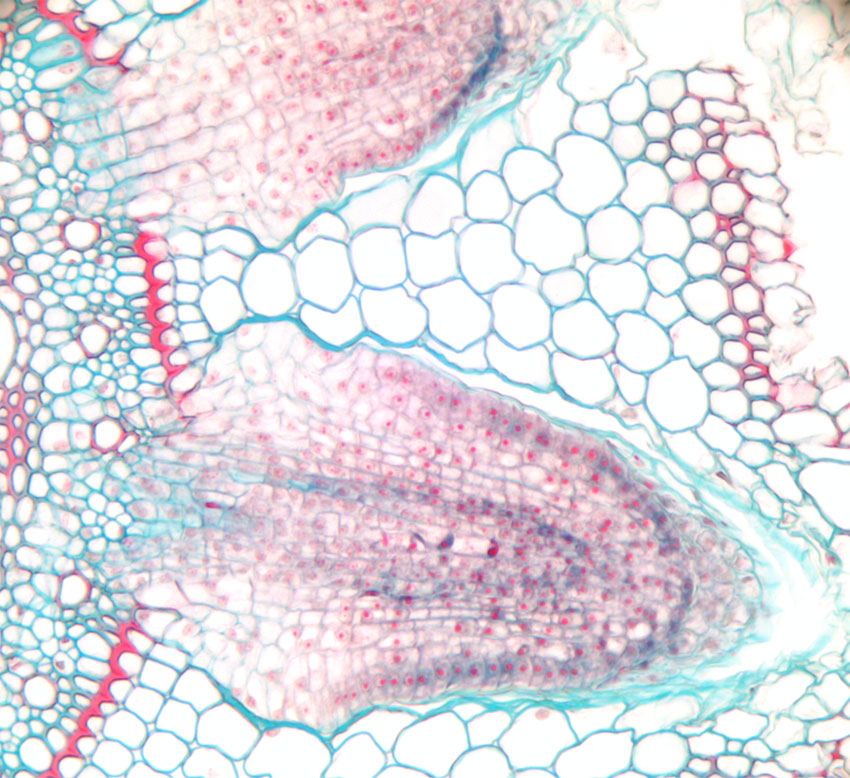
Bilde 2. Utsnitt av rot av mais (Zea mays) som viser en siderot som utvikles fra pericykel og som trenger seg gjennom rotbarken (cortex).
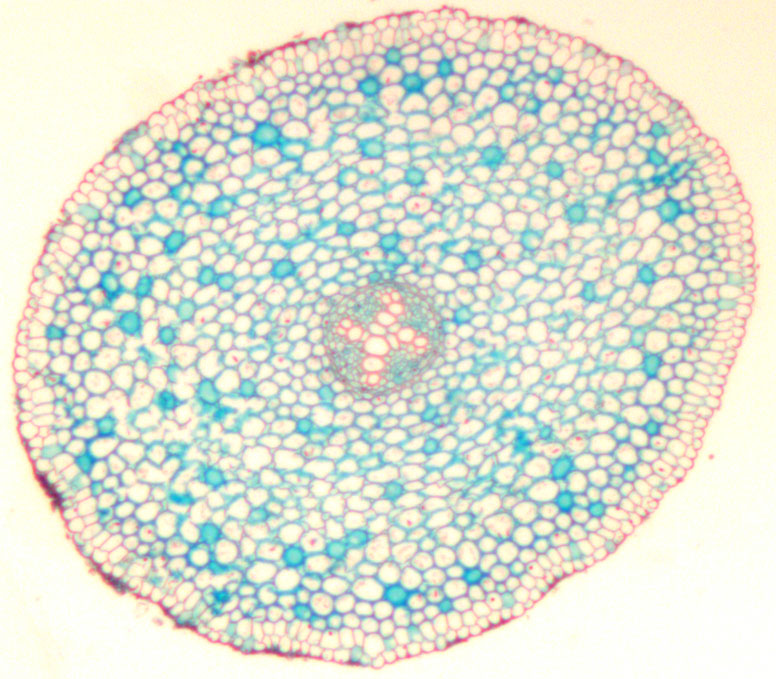
Bilde 3. Tverrsnitt av rot fra krypsoleie (Ranunculus repens L.).
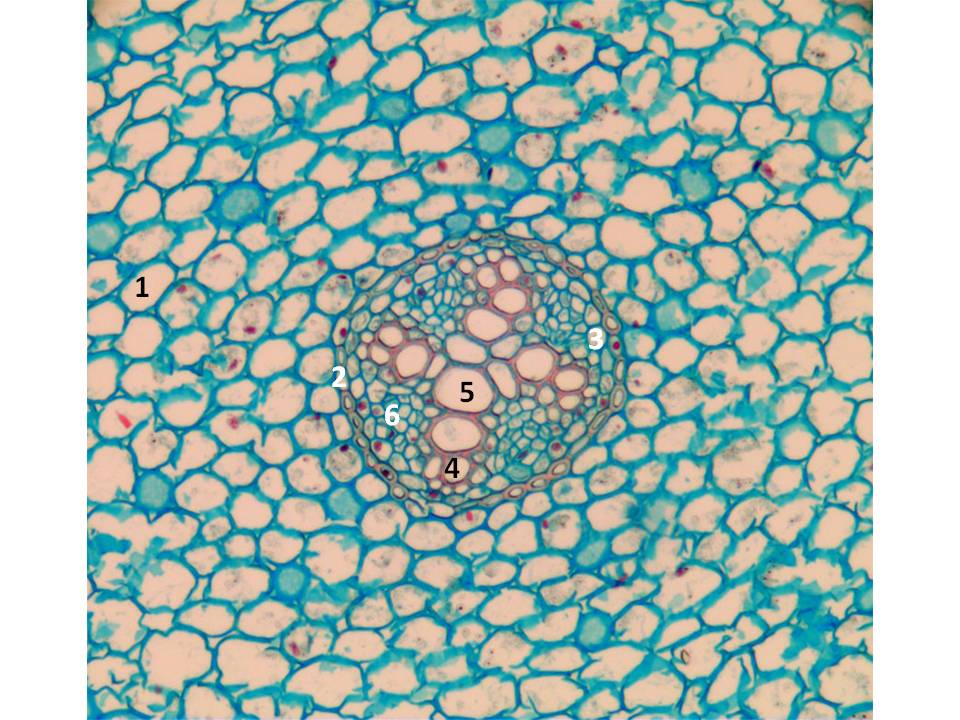
Bilde 4. Tverrsnitt av rot fra krypsoleie (Ranunculus repens L.). 1: Rotbark (cortex). 2: Endodermis. 3: Pericykel. 4: Protoxylem. 5: Metaxylem. 6: Primært floem. Både protoxylem og metaxylem er primære vev dannet fra det rotapikale meristemet. Etter hvert som rota blir eldre vil det bli dannet et vaskulært kambium mellom xylem og floem, og dette kambiet vil laget sekundært xylem og sekundært floem og rota øker i tykkelse. På eldre røtter slites vekk deler av cortex, og det blir dannet et korkkambium som lager periderm. Protoxylemet ligger ytterst i de fire armene med xylem.
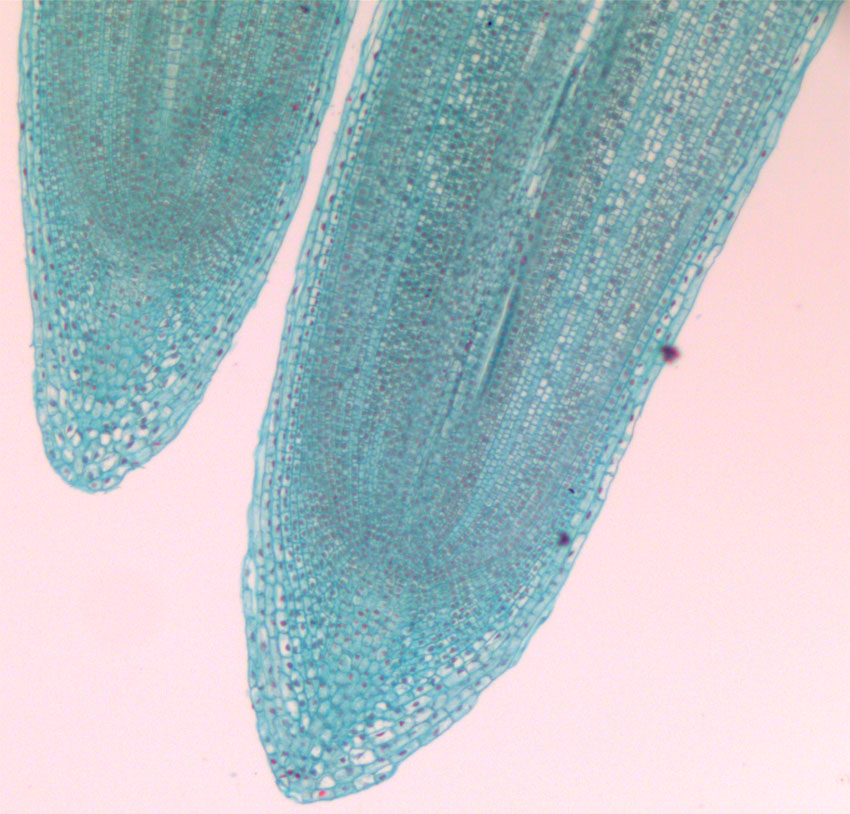
Bilde 5. Lengdesnitt av rotspiss fra løk (Allium sp.). Det apikale rotmeristemet er dekket av en beskyttende rothett med en sentralt plassert kolumella.

Bilde 6. Lengdesnitt av rotspiss fra løk (Allium sp.). Detalj fra overgangen mellom rothette og rotapikalt meristem.

Lengdesnit av rotspiss av løk. nederst vises rothetta med celler som fornyer seg og beskytter det rotapikale meristemet. Rothetta skiller ut karbohydratslim som gir vekst av bakterier og gjør også at rota glir lettere gjennom jordsmonnet.
Rotknoll
En rotknoll er et underjoridsk lagringsorganer i planter laget fra omdannete siderøtter som svulmer opp og blir fylt med opplagsnæring, vesentlig i form av stivelse og er derfor morfologisk forskjellig fra stengelknoller er omdannete stengler. Til forskjell fra en rotstokk som er en underjordisk stengel. Eksempler på rotknoller er kassava (Manihot esculenta), søtpotet (Ipomoea batatas) og yamsrot (Dioscorea sp.).
Rotarkitektur
Helt fra Devon, 360-400 millioner år siden, hvor planter kunne overleve på land ble det utviklet rhizoider og røtter som kunne trenge igjennom jorda og ta opp vann og mineralnæring. Røttene danner et komplekst 3-dimensjonalt topologisk nettverk hvis form og struktur blir påvirket av ytre miljøbetingelser. Rotdybde, rotoverflate, og grad av greining blir påvirket av vanninnhold, saltinnhold, oksygenkonsentrasjon og spesielt av makronæringsstoffene nitrogen og fosfat. Rotsystemet danner en kobling mellom jord og atmosfære, og påvirker den delen av jorden som ligger nærmest røttene, rhizosfæren. Plantene kan påvirke pH i rotsonen, hvor mineralnæring blir tatt opp via ionebytting fra jordkolloidene. F.eks. ved jernmangel kan en plante surgjøre rotsonen, noe som øker tilgangen på jern. Røttene skiller ut karbohydrater og andre kjemiske stoffer fra rothetta og rotspiss som gir opphav et spesielt mikrobiom i rhizosfæren. Røttene blir omgitt av en rhizoskjede med jordpartikler, og påvirker jordstrukturen. Døde planterøtter danner et reservelager av organisk materiale i jorden. I interaksjon med sopphyfer blir det dannet ektomykorhiza eller endomykorhiza. Tyngdekraften påvirker rotveksten via gravitropisme. Ved rothalsen, i overgangen mellom rot og stengel skjer det en endring i plasseringen av ledningsvevet fra sentrum i rota over til periferi av stengelen.
Via signalpeptider som CLAVATA 3 (CLV3), C-terminal kodet peptid (CEP1) og ESR-relatert peptid (CLE) blir graden av siderotdannelse påvirket.
Radikal (l. radix) - gå til roten.