Forskjellige former for pollinering
Pollinering kan skje ved:
1) Vindpollinering (anemofili, gr. anemos - vind; philein - å elske) hvor blomsten er uanselig, åpent plassert, mangler duft og nektar (gras, rakletrær) og hvor lett pollen spres i store mengder, ofte tidlig om våren. Nåletrær har pollen med luftsekker.
2) Insektspollinering (entomofili, gr. entomen - insekt) hvor blomstene er store iøyenfallende, har farget blomsterdekke med saftmerker og ofte zygomorfe blomster hvor insektet krabber inn, blir dekket av pollen som blir deretter overtørt til neste blomst (chasmogami). Insekter flyr eller forflytter seg fra blomst til blomst og tar med seg pollen som gir krysspollinering av blomstene. Blomsten kan ha spesielle utskillende organer, nektarier, som produserer nektar med sukker og aminosyrer som er vktige for insektene. Pollen har pollenkitt og ytre struktur som gjør at de lett festes fast. Pollen er næringsrikt og inneholder protein. Hos humler kan arbeidshumlene spesialisere seg på enten innsamling av nektar eller pollen. Hos både humler, veps og bier blir pollen gitt som næring til larvene. Humler har en lang tunge som gjør at de kan suge nektar fra planter med lange kronrør, e.g. kløver.

Insektpollinering av gressløk
Dessuten produserer humlene egenvarme som gjør at de kan fly i kaldt og fuktig vær om våren hvor de er viktige pollinatorer under slike værforhold.
Sommerfugler har en lang opprullet sugesnabel som suger nektar.: Blomsten ofte tallerkenform med langt tynt kronrør, for eksempel nellik.

Neslesommerfugl (Nymphalis urticae) på blomst av vanlig gressløk.
Nattsvermere: Lysende hvite blomster med sterk duft (nattfiol, nikkesmelle). Åtselinsektpollinering: Brune, grønne og grå blomster, ofte med åtselduft.

Svalestjert på tistel.
Noen planter har sterkt spesialiserte artstro insekter som sørger for pollinering, spesielt velutiklet for arter i orkidefamilien, og er et resultat av koevolusjon.
3) Fuglepollinering spesielt i tropene (ornithofili, gr. ornis - fugl; philein - å elske): Store blomster med grelle farger, spesielt knallrød (kolibrier i Amerika, honningfugl i Afrika).
4) Flaggermuspollinering (chiropterofili, gr. cheir - hånd, pteron - ving): Store kraftige hvite-grønlige blomster utenfor bladverket med rikelig nektar.
5) Pollinering kan også skje via vann (hygrofili) hvor blomstene er uanseelige uten skiltapparat. Hannblomstene kan også løsne og flyte på vannet f.eks. ålegras, eller andemat.
Pollineringen kan også deles i
A. Selvpollinering (autogami, gr. autos - selv; gamos - ekteskap) hvor pollen overføres til arr innen samme blomst. Blomster som selvpollineres kan også i visse tilfeller krysspollineres. Det finnes også mekanismer som fremmer selvpollinering med blomster som ikke åpner seg (kleistogami gr. kleistos - lukket, skjult) hvor genotypen beholdes i stabilt miljø (fiol).
Nabopollinering (geitonogami, gr. geiton - nabo) som skjer mellom forskjellige blomster på samme plante.
B. Krysspollinering (fremmedpollinering) (xenogami, gr. xenos - fremmed) som skjer mellom blomster på forskjellige planter. Dette siste sikrer genetisk rekombinasjon og gir evolusjonsfordeler. Det er derfor mange mekanismer som fremmer sannsynligheten for fremmedpollinering.
Hvordan hindre selvpollinering
Plantene har en rekke mekanismer som hindrer selvpollinering. Selvpollinering gir økt homozygoti og derav mindre genetisk variasjon, sammenlignet med krysspollinering som fremmer dannelse av heterozygoter med økt variasjon som evolusjonen kan virke på. Mekanismer som gir størst sannsynlighet for å hindre selvpollinering er:
1) Selvinkompatibilitet (selvsterilitet, selvuforenlighet) hvor plantens eget pollen ikke vil spire på arret. Selvinkompatibilitet SS styrt av genprodukter fra et S-lokus, et gen uttrykt i den hannlige gametofytten og det andre uttrykkes i griffel. Selvinkopatbilitet girS-lokusproteiner i den hannlige gametofytten interagerer med S-lokusprotiner på arr, griffel eller embryosekk. Hvis de tilhører samme haplotype starter en prosess med å hindre befruktning. Hos gras er det et flerlokus system.
Prezygotisk selvinkompatiblitet , hvor pollen forkastes og avvises på grenseflaten arr, griffel eller frøemne er den eldste formen for selvinkompatbilitet
2) Heterostyli hvor blomstene har forskjellig lengde på griffel hos individer av samme art slik at pollenknappene blir sittende i forskjellig høyde i forholdet til arret (dimorf heterostyli hos nøkleblom, Forsythia, lungeurt, bukkeblad; trimorf heterostyli hos kattehale). Heterostyli gir blomster med 2-3 morfologiske typer basert på lengden av griffel og pollenbærer
3) Pollen og arr kan modnes til forskjellig tid (dikogami gr. dicha - atskilt). Via førsthannlighet (protandri, gr. protos - først; aner - han, førsthanlig) slik som blomsten hos geitrams, eller førsthunnlighet (protogyni, gr. protos - først; gyne - kvinne, førsthunnlig) hos kjempe og hårfrytle. Planter som har feller som fanger insekter for en kortere periode før de slippes fri er ofte førsthunnlig. Pollensekkene og arr kan også ha en romlig plassering slik at selvpollinering hindres eller hemmes (herkogami gr. herkos - hinder) som hos orkidéer og sverdlilje.
4) Hannsterilitet hvor planten har mistet evnen til å lage funksjonelt pollen
Pollenspiring
Prezygotisk krysningsgjenkjenning hvor pollen fester seg og hydratiseres på arroverflaten og pollenlangen må vokse fram til mikropylen på frøemne.Arret er dekket av celler med papiller hvor pollen kan feste seg. Avhengig av art er det tørt eller vått arr. Tørt arr har en overflate med tørre cellevegger, dekket av kutikula og en proteinpellikel. Vått arr er dekket av en klebrig væske bestående av protein, polysakkarider og fett. Etter at kompatibelt pollen har festet seg på arret skjer det en hydrering av pollen via kapillart vann. Ved hydreringen lages deg en kalsiumgradient som gir en polarisering av pollenkornet før pollenslangen vokser ut. Pollenslangen vokser gjennom epidermis i arret. Der hvor pollenslangen vokser ut er det høy konsentrasjon av kalsium (Ca2+), og aktinfilamenter og sekretoriske vesikler frakter celleveggmateriale til spissvekst av pollenslangen. GTPase som hydrolyserer GTP til GDP bidrar til å regulere polaritet i cellen. I griffelen er det et ekstracellulært matriks med prolin og hydroksyprolinrike glykoproteiner og arabinogalaktanproteiner. Veksten av pollenslangen via transmisjonsvev kan forklares med en mekanohypotese eller en kjemotropisk hypotese. Arr og griffel inneholder cysteinrike adhesiner (lipidoverføringsproteiner). Kemocyanin er et lite basisk kobberprotein funnet i griffelen hos liljer og som deltar i veksten av pollenslangen. Pollenslangen vokser vanligvis gjennom mikropylen til frøemnet og avleverer to spermceller til den hunnlige gametofytten. Pollenslangen slutter å vokse, spissen brister og frigir de to spermcellene.
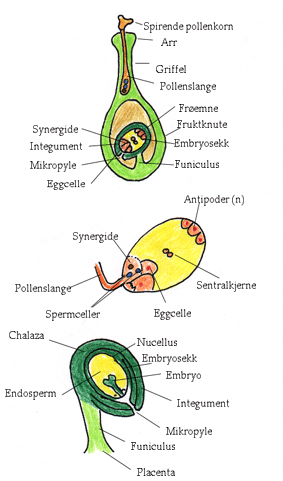
Kompatibelt pollen lander på arret, tar opp vann og det vokser ut en pollenslange som vokser gjennom transmisjonsvev i griffelen fram til mikropylen i frøemnet, hvor en av spermkjernene i spermcellene fusjonerer med egget og den andre med de to sentralkjernene i sentralcellen og danner triploid endosperm. Zygoten danner et embryo under embryogenesen, hvor spissen på kimroten peker mot mikropylen
Selvinkompatibilitet (selvuforenelighet)
For gametofyttselvinkompatibilitet vil pollenkornet spire på arret hvis den haploide genotypen til pollen er forskjellig fra S-lokus på det hunnlige arret. For ikke-kompatibelt pollen blir RNA i pollenslangen nedbrutt av cytotoksisk RNase i den øverste delen av griffelen eller via programmert kontrollert celledød. Barriærene som hindrer selvpollinering ved selvinkompatibilitet kan befinne seg ved arret, i griffel, eller ved inngangen til embryosekken.
Postzygotisk selvsterilitet skjer etter at det er dannet en zygote, men hvor zygoten og/eller endospermen stopper å utvikle seg.
Sporofyttinkompatibilitet skyldes rester av sporofytten blir sittende igjen på utsiden av pollenkornet og blir bestemt om den diploide genotypen til pollen er forenlig med genotypen til hunnen. Ved sporofyttinkompatibilitet blir ikke pollenkornet hydrert, og signaler via reseptorkinaser og peptider hindrer pollenslangen i å vokse.
Pollinering og befruktning
Pollinering er overføring av pollen fra pollenbærer til arr. Pollineringen skjer med vind, insekter, dyr og fugler. Det er to hovedtyper arr: tørre arr og våte arr.
Våte arr f.eks. hos løvemunn og liljer består av celler som produserer glykoproteiner, slim og næring som er nødvendig for at pollenkornet skal spire. Tørre arr er evolusjonsmessig mer avansert og finnes hos gras, kål og trær. Griffelen kan være åpen eller lukket. En åpen griffel har et hulrom i midten med indre epitelceller dekket med polysakkarider, lipoproteiner, og glykoproteiner. En lukket griffel har et transmitterende vev som leder pollenslangen. Pollenslangen vokser i spissen. Veggen i pollenslangen inneholder ikke cellulose, men kallose som er et β-1,3-glukan. Kallose lages i Golgiapparatet og transporteres i form av vesikler til spissen av pollenslangen. Det er høy konsentrasjon av kalsium (Ca2+) i spissen av pollenslangen.
De fleste planter er selvsterile, det vil si at eget pollen ikke vil spire på eget arr, en egenskap som fremmer krysspollinering. Gener som bestemmer cytoplasmatisk hannsterilitet finnes i mitokondriene.Planter med genet cms har ikke levedyktig pollen, og egenskapen nedarves maternalt via mitokondriene. Programmert celledød kan gi prematur aldring av pollenknappen, og mitokondrier kan delta i programmert kontrollert celledød.
Gametofytt selvinkompatibilitet skyldes at et genprodukt fra den hannlige gamtofytten reagerer med et genprodukt i det hunnlige vevet i griffelen. Interaksjonen kontrollert av et S-locus som har titalls forskjellige alleler i en naturlig populasjon. Hvis S-allelet er likt i pollen og hunnlig del vil ikke pollenslangen vokse og utvikle seg. Proteinproduktene fra S-allelet er basiske glykoproteiner eller enzymet RNase som spalter RNA. Ved sporofott selvinkompatibilitet er det alleler i rester av sporofytten som sitter igjen på pollenkornet som deltar. Kompatibelt pollen, som den hunnlige delen godtar, tar raskt opp vann og pollenslangen begynner å vokse etter kort tid. Det skjer en dobbel befruktning (syngami) ved at en spermcelle fusjonerer med egget og gir zygote. Den andre spermcellen fusjonerer med to polkjerner og gir triploid endosperm. Både spermceller, eggcelle og sentralcelle er nakne protoplaster.
Hannsterile planter er mer kostnadsbesparende enn manuell fjerningen av toppen av plantene for å hindre selvpollinering i produksjonen av maishybrider. Tidligere ble hybridmais med cytoplasmatisk hannsteriltet type cmsT (T for Texas) benyttet og dette genet laget et URF13 (13 kilodalton) som gir hannsterilitet. Det oppstod en sykdom (“Southern corn blight”) som ødela store deler av hybridmaisavlingene. Dette skyldtes at soppen Bipolaris maydis syn Cochliobolus heterostrophus lager et HmT toksin som reagerer med URF13 og lager porer i den indre mitokondriemembranen. Nå benyttes genmodifiserte hannsterile planter basert på genene som koder for barstar og barnase.
Den hannlige gameten overleveres til frøemnet gjennom en rasktvoksende pollenslange fra det haploide pollenkornet. Pollen består av en stor vegetativ celle og en generativ celle eller to spermceller. Spermcellene lages fra mitotisk deling av den generative cellen. Når pollenkornet lander på arret lager den vegetative cellen en utvekst som blir pollenslangen som vokser gjennom det transmitterende vevet i griffelen ned til frøemnet. Pollenslangen avleverer de to spermcellene i embryosekken hvor de smelter sammen med henholdsvis eggcellen og sentralcellen. Veksten av pollenslangen er avhengig av et cytoskjelett av aktin.
Profilin er et protein som binder seg til globulært aktin i pollenslangen og virker som et bindeledd mellom aktin og fosfatidylinositol. I noen tilfeller har flavonoler på utsiden av pollenkornet betydning for vekst av pollenslagen