Vannmangel i land- og hagebruk vil gi betydelige reduksjon av avlingspotensialet. Store deler av Jordens ferskvannsressurser blir benyttet til kunstig vanning av landbruksvekster (irrigasjon), noe som resulterer i synkende grunnvannstand, og økt salinisering av jorden med tilhørende saltstress. Store mengder vann fordamper fra jorden via plantene, noe som gir et bedre mikroklima å puste i under trær eller på en grønn gressplen, sammenlignet med en balkong i betong eller treplatting hvor det ikke er slik fordampning av vann. Om vinteren kan frossen jord gi vintertørke. Om sommeren er det ofte høy temperatur sammen med tørke, noe som kan gi varmestress.
Vann til avkjøling, transport, styrke og fotosyntese
Mesteparten av vannet plantene tar opp fra jorda via røttene brukes til temperaturregulering og avkjøling bladverket, til transport av mineralnæring, og til opprettholdelse av turgortrykket i cellene, mens bare 1-2% av vannet som blir tatt opp brukes til fotosyntese. Det er røttene som først registrerer mangel på vann, og gir signal om dette til skuddet via plantehormonet abscisinsyre. Hvis fotosyntesen i flaggbladet hos kornslagene blir redusert på grunn av vannmangel, blir mengden korn redusert, siden flaggbladet har en viktig funksjon i transport av fotosynteseprodukter til kornakset. Veksten av røttene er avhengig av fotosynteseprodukter fra bladene i de overjordiske delene. Synker vann-nivået i jorden raskere enn rotveksten kan planten visne ved permanent visningspunkt. Plantene har forskjellig evne til å utnytte vannet (WUE).
Tilpasning til tørke ved å redusere vanntap
Plantenes første respons på vannmangel er å lukke spalteåpninger i blader og grønne stengler. Plantene tar opp karbondioksid (CO2) som trengs i fotosyntesen gjennom spalteåpningningene, men samtidig tapes store mengder vann gjennom de samme åpningene. Planter har en avansert spalteåpningsmekanisme som regulerer vanntapet. Skog og annen vegetasjon spiller en stor rolle i vannsyklus på Jorden. Plantene bruker fra 300 liter til 1000 liter vann for å kunne produsere 1 kilo biomasse. Det betyr at hvis et land som har mangel på vann importerer matkorn importerer det samtidig vann.
Selv om spalteåpningene utgjør bare ca. 1-2% av bladarealet tapes ca. 90% av vannet gjennom disse, mens det resterende vannet tapes gjennom bladoverflaten dekket av celler i en epidermis med vanntapsbeskyttende kutikula, og et lag med voks. Planter er helt avhengig av vann for å kunne vokse, slik at ved vannmangel stopper veksten med tilhørende veksttap. Hos urtaktige planter og i blader har vannet også en viktig funksjon i opprettholdelse av styrke sammen med styrkevevet, men når vanntrykket i cellene synker blir bladene hengende, skrumper og krøller seg. Gras visner raskt ved tørke, men gjenoppretter veksten raskt hvis det blir god vanntilgang. Arter som ryllik og balderbrå har velutviklet rotsystem og kan motstå tørke. Hagejordbær har et grunt rotsystem og er tørkeutsatt.
Tørke kan forekomme sammen med høy temperatur og gi varmestress.

Tørke av blåbærlyng sommeren 2018. Brunfargen skyldes oksidasjon av fenoler og garvestoffer.

Tørkeskader på blad av spisslønn sommeren 2018. Mest tørkeskader og nekrose langs bladranden, men hvor det indre av bladet skades i mindre grad.
Tilpasninger til lite vann styrt av plantehormoner og signalveier
Plantene har stor evne til å tilpasse seg endringer i vekstbetingelsene, en adapsjon, styrt via molekylære signalveier og signaloverføring i cellene. Det omfatter økt biosyntese av kutikula og voks på bladene, økt vekst av røttene på bekostning av skuddet, rotvekst mot deler av jorden som inneholder mest vann, samt avkasting av blader, som kan sees som blader som får gulfarge eller gule gressplener. Slik tilpasning hos flerårige vekster vil gi ny vekst hvis plantene får ny tilgang på vann. Vanning av gressplen er misbruk av vann, bortsett fra i spesielle tilfeller. Mer følsomme for vannmangel er sommerblomster, planter med et grunt rotsystem slik som hagejordbær, samt ettårige landbruksvekster.
Mange av tilpasningsresponsene er styrt via plantehormonet abscisinsyre, som nevnt tidligere. Kaktus og andre sukkulenter som vokser i tørre områder i ørken og sklerofyllvegetasjon i i områder sommertørke og middelhavsklima har i utgangspunktet en rekke tilpasninger til lite vann. Oliventrær og oleander som blomstrer langs veiene er eksempler på tørketilpassete xerofytter.

Oleander (Nerium oleander) i gravmyrtfamilien (Apocynaceae) er eviggrønn busk med tørketilpassete blad. Danner kratt langs veier i middelhavsområdet. Finnes i former med hvite, rosa eller rød blomster. Oleander er giftig og inneholder hjerteglykosidet oleandrin. Brukes også som stueplante.
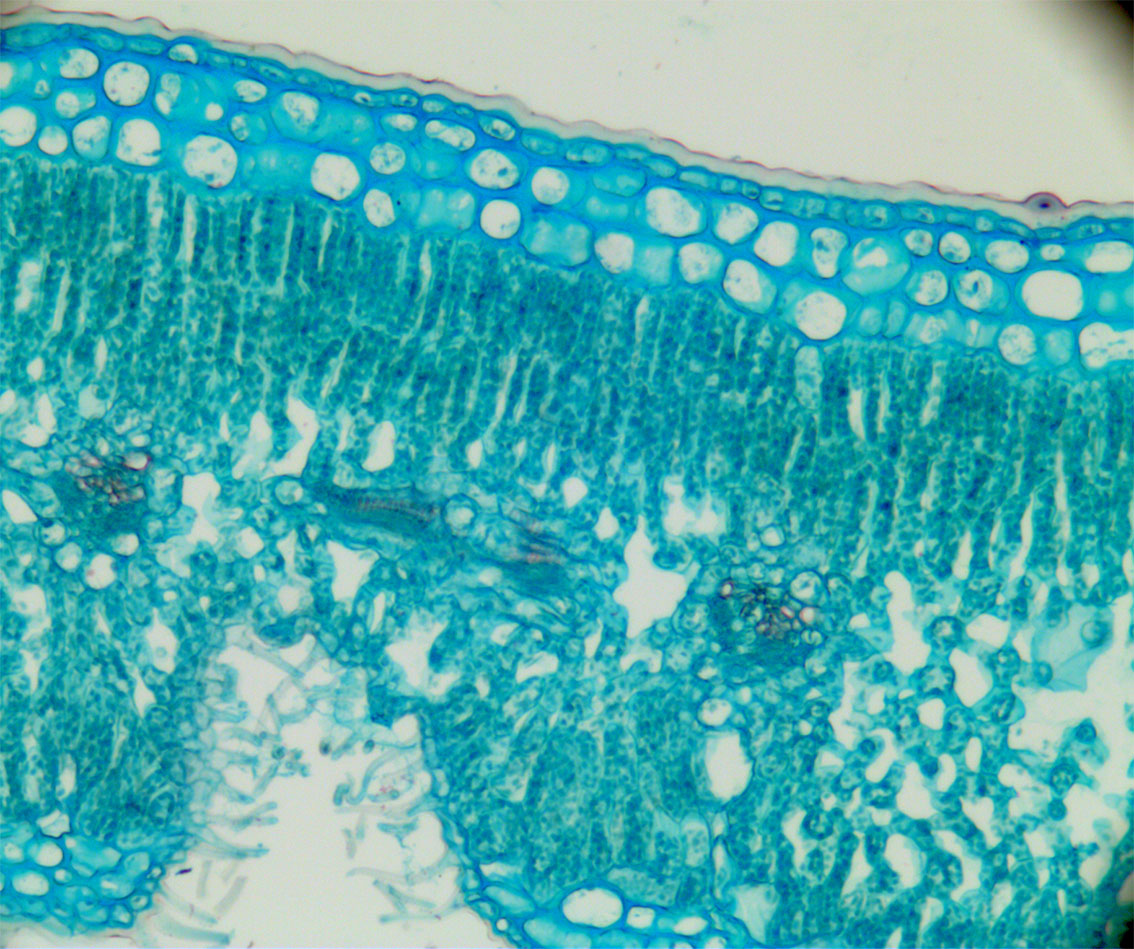
Tverrsnitt av blad fra oleander (Nerium oleander) som vser tilpasning til tørke. spalteåpningene ligger nedsenket i et hulrom fra undersiden av overflaten til bladet og omgitt av kjertelhår som øker tykkelsen på grenselaget. Oversiden av bladet er dekket av et tykt vokslag og under dette epidermis med tykke cellevegger og under denne ytterligere to lag med tykkveggete celler (hypodermis) før man kommer til de fotosyntetiserende cellene i palisadevevet med kloroplaster. Svampvevet med mer hulrom mellom cellene ligger under palisadevevet.
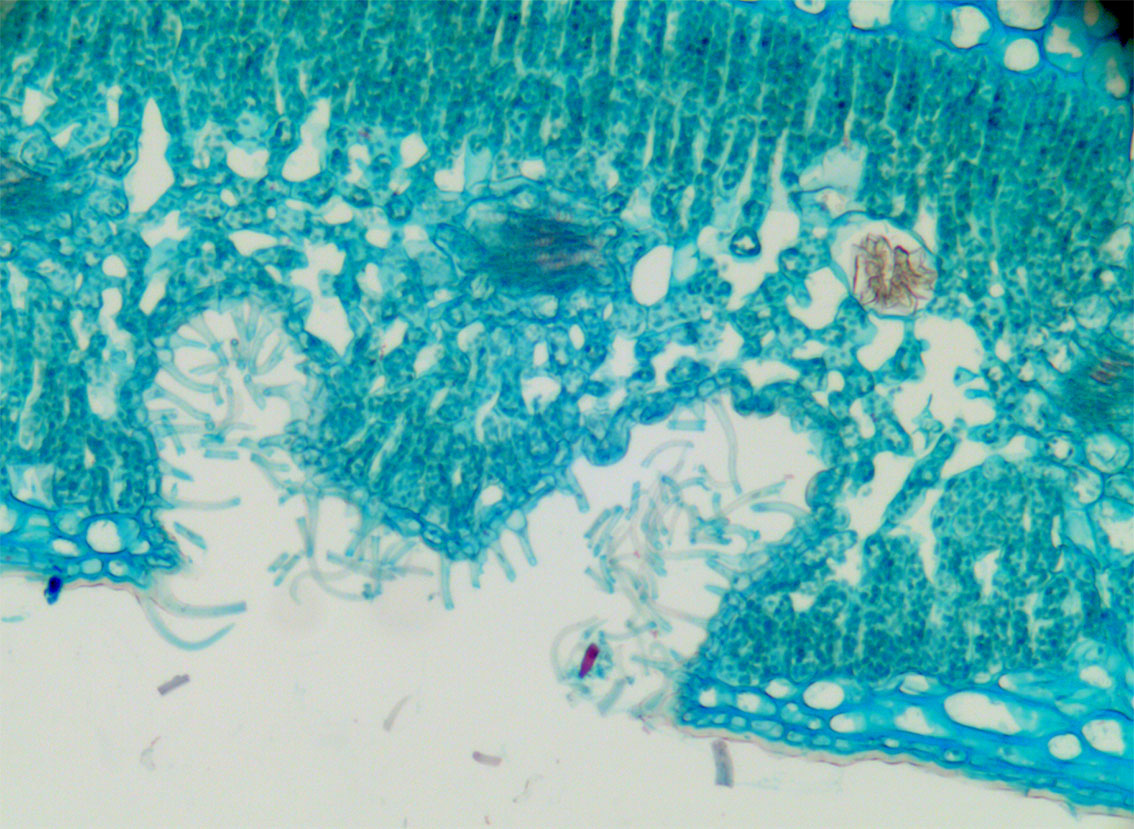
Tørketilpasning i bladet fra Nerium oleander som viser to hulrom i undersiden på bladet hvor de i bunnen av hulrommet er spalteåpninger. Hulrommet er fylt med trikomer som gir luftstille rom rundt spalteåpningene og som reduserer transpirasjonen.
Tørketilpasning kan være en type metabolisme og fotosyntese som gir redusert behov for å ha spalteåpningene åpne om dagen, som hos de såkalte CAM-planter og C4-planter. Disse har høy vannutnyttelseseffektivitet. En annen vekststrategi benyttes av efemerer. Imidlertid, planter kan dø ved tørke og vannmangel hvis de når et permanent visningspunkt.
Det er rota som registrer at planten er utsatt for vannmangel. Det kan gi seg utslag i en rekke effekter på planten. Minsket syntese av cytokininer og økt syntese av abscisinsyre i røttene kan gi bladaldring og avkastning av blad. Planten kan endre bladenes fysiske orientering i forhold til sollyset slik at de mottar minst mulig lysenergi. Er plantene utsatt for tørke over lenger tid vil det gi økt mengde voks på overflaten av kutikula, og mengden av spalteåpninger på bladene kan reduseres. Tørken kan gi spesielle knollformete røtter uten rothår, som seinere kan gi opphav til nye røtter når vanntilgangen bedres. Økning i innholdet av abscisinsyre vil gi lukking av spalteåpninger. Abscisinsyre kan også gi økt sukkertransport ut av bladene, som som igjen gir mer negativt vannpotensial i bladcellene slik at de lettere kan få tak i vann. Abscisinsyre øker den hydrauliske konduktiviteten for vann i røttene og øker transporten av ioner inn i xylemet. Abscisinsyre blokkerer samtidig utskillelsen av protoner fra kloroplastene. Under tørken kan det skje kavitering i ledningsvevet, noe som vil redusere vanntransporten og ytterligere forverre situasjonen for planten eller treet. Ved tørke forkortes ofte lyssyklus til planten.
Det er rota som er den som registrerer vannmangel først. Tørke resulterer i:
☞ Bladaldring og felling av blad.
☞ Økt syntese av epikutikulær voks.
☞ Endret bladorientering.
☞ Minsket stomatakonduktanse.
☞ Rask refordeling av rotbiomasse.
☞ Kort livssyklus for planter tilpasset strøk med lang tørketid.
Ved tørke lages spesielle røtter uten rothår, som seinere kan gi opphav til nye røtter. Når vann beveger seg fra celle til celle kan det følge akvaporiner med høy hydraulisk konduktanse eller gjennom lipidlag med lav permeabilitet. Fosforylerte akvaporiner er åpne, ikke-fosforylerte er lukket. Ved tørke kan akvaporiner lukkes.
Tørke gir redusert frøspiring
Når frø skal spire er den første fasen vannopptak (imbibering). I tillegg får den unge frøplanten vanskeligheter med å komme i kontakt med jord med høyt nok vanninnhold til at veksten kan fortsette. Tilsammen gir dette redusert spireprosent.
Tørke, erosjon, jordstruktur og sprekkdannelse
Jord har tre faser: fast fase, vannvase og luftfase. Tørke og evapotranspirasjon (fordampning målt med et lysimeter) påvirker derved strukturen i jorden. Det blir dannet nye former for jordpartikkel- og kolloidaggregater. Jordoverflaten får sprekkdannelser med et karakteristisk fure- og sprekkmønster med polygoner (tesselering) når jorden skrumper inn, spesielt i leirrik jord (vertisol). Leirpartiklene, som er sjiktformete aluminiumsilikater), kan både absorbere vann, ekspandere og svelle, eller avgi vann og skrumpe. Når de tørker trekker de seg sammen, danner aggregater og leirklumper. Når nedbøren endelig kommer har jorden vanskeligheter med å ta opp vannet, men vann får økt mulighet til å trenge dypere ned i jorden via sprekkene, og ikke bare renne vekk fra den hard jordoverflaten. Imidlertid, er sprekkene dype kan planterøttene miste muligheten til å utnytte vannet.

Leire og leirjord får dype sprekker ved tørke.
Tørr jord er dessuten utsatt for vinderosjon og tap av jord, siden røttene fra den tørre døde vegetasjonen mister evnen til å holde på jorden.
Tørke gir redusert vannpotensial i jorden, senker hydraulisk konduktivitet (Darcys lov), og vannet blir derved mindre tilgjengelig for plantene.
Akvifer
Senking av grunnvannstand ved tørke kan via påvirkning av jordstrukturen gi synking og sprekker i bygningskonstruksjoner som er bygd på jordoverflaten. Å fylle et grunnvannsreservoir, et akvifer (l. aqua - vann; ferre - bære, føre) bestående av porefylt berggrunn, grus og sand, kan ta opptil tusen år, eksempel i California og Midtvesten i USA, men det kan bli tømt på relativt kort tid via stadig dypere brønnboring for å sikre vann til matproduksjon (irriasjon). En akvifer kan få tilførsel av vann via nedbør, elver og vann, men kan også bli forurenset ved inntrengning av urent vann eller saltvann. Det kan være forskjellig hydraulisk konduktivitet i deler av akviferen, og forskjellig porøsitet og permeabilitet i bergrunnen som påvirker vannfluksen (Darcys lov). Guarani akvifer (Sør-Amerika), Ogallala akvifer (USA), Edwards akvifer i Texas, det store artesiske bassen (GAB, Australia), den nubiske sandsteinsakviferen (N-Afrika) er eksempler på store akviferer. Hadde vi lagt Gardermoen flyplass et annet sted, kunne Osloregionen hatt en akvifer på romerikssletta. I motsetning til overflatevann inneholder ikke sandfiltrert vann tarmparasitter.
Tørkestress
Et grunt rotsystem er følsomt for uttørking, men kan respondere raskt på nedbør. Et dypt rotsystem kan gå ned til en mer stabil vanntilgang. Vannmangel øker mengden voks utskilt på bladoverflaten. Tørkestress og varmestress er koblet med hverandre. Redusert bladstørrelse og bladorientering kan gi vannsparing. Bladene kan rulles sammen for å redusere den eksponerte overflaten f.eks. gras. Visning gjør at bladene henger ned og mottar redusert innstråling. Hydropassiv lukking av spalteåpningene for å redusere vanntap skjer ved at vann fordamper direkte fra lukkecellene ut i atmosfæren. Hydroaktiv lukking av spalteåpningene skjer ved at røtter og skudd dehydreres, ioner fraktes ut av lukkecellene etterfulgt av redusert turgor i lukkecellene. Abscisinsyre gir lukking av spalteåpningene ved vannmangel.
Plantene kan ta opp vann så lenge som vannpotensialet i plantene er lavere enn vannpotensialet i jorda. Et mildt tørkestress reduserer fotosyntese og stomatakonduktanse. Cellene og røttene skrumper (cellulær dehydrering) ved vanntap og rothårene skades. Når jorda tørker ut minsker vannkonduktansen inntil permanent visningspunkt ved ca. -1.5 MPa hvor planten ikke lenger kan gjenskape turgortrykk og rehydrere planten. Ved tørke minsker denhydrauliske konduktivitet, kollaps av cellevegger gir cytorhysis og røttene skrumper. Ved tørke blir den ytre del av cortex i rota, kalt hypodermis (et cellelag under epidermis), dekket av suberin som minsker vanntapet.
Vanligvis er det osmotiske potensialet i planteceller fra en velvannet plante fra -0.8 til 1.2 MPa. Kavitering skjer ved -1 til -2 MPa. Vanndefisit vil si at vanninnhold i celler eller vev er lavere enn det høyeste vanninnholdet i fullhydratisert vev. Vanndefisit minsker vannkonduktansen i planten. Et tilstrekkelig turgortrykk er nødvendig for vekst av celler.
\(vekstrate = m\left(\Psi_p-Y\right)\)
hvor Y er turgortrykket som må overstiges for at celleveggen skal kunne ekspandere, Ψp er trykkpotensialet i cellen (turgortrykket), og m er celleveggekstensibilitet dvs. evne til å respondere på trykk, og består av en elastisk og en plastisk komponent. Vanligvis er Y=0.1 - 0.2 MPa.
Osmotisk tilpasning opprettholder vannbalansen. Tørkestress gir akkumulering av oppløste stoffer kalt osmolyter (prolin, betain, glycinbetain, mannitol, sorbitol) i cytoplasma. Ved å akkumulere osmolyter kan cellene opprettholde turgortrykket som er nødvendig for vekst. Reduksjon i vanntilgangen reduserer veksten av skuddet og øker veksten av rota, og det skjer en fysiologisk adaptering ved bladavkasting å redusere bladarealet i forhold til røttene, hvor abscisinsyre deltar. Noen planter kaster av alt bladverket før tørkesesongen begynner, via nysyntese av etylen. Lite vann bestyr at planten må investere mer i røtter og i oppløste stoffer på bekostning av skuddvekst. Røttene vokser så lenge som fotosyntesen i skuddet gir nok fotosynteseprodukter til røttene. Ved vannmangel blir fotosynteseprodukter også kanalisert til frø og frukt. Fotosyntesen er mindre følsom for vannmangel enn veksten av blad. Osmotisk potensial fra -1 til -2 MPa er et mildt vannstress, men fra -2 til -5 MPa er et resultat av betydelig tørkestress. Celler blir flakkide og planten visner når turgortrykket går mot null. Halofytter har lavt osmotisk potensial.
Tørketoleranse er plantenes evne til å tåle dehydrering. C4- og CAM-planter har en fotosyntesemetabolisme tilpasset lite vann, og utnytter ressurser i tørre områder maksimalt. Osmotisk stress og tørkestress kan indusere CAM-metabolisme hos planter som kan skifte mellom C3 og CAM avhengig av vannforholdene. Tørkeunngåere tilpasser veksten i den perioden vann er tilgjengelig, f.eks. regntid eller annen vekstgunstig årstid. Tørkeutsettere har evne til å opprettholde hydratisert vev f.eks. kassava (Manihot esculenta) og cowpea (Vigna unguiculata).
Osmotisk stress gir endring i genekspresjon av stresskontrollerte gener med økt syntese av vannkanalprotein, varmesjokkproteiner, og enzymer som lager osmolyter,
Mer om vannsyklus.